Построение моделей особей медоносных пчёл
и их информационных связей жизнедеятельности © Ю.Н.Третьяко

8. Определение пола и половые акты у родителей пчел.8.1. Эволюция систем определения пола медоносных пчёлЭволюция систем определения пола означает постепенный закономерный переход поколений пчёл от одного полового состояния в другое регулируемое естественным отбором. Пол – это совокупность свойств организма, обеспечивающих функцию воспроизводства потомства и передачу генетической информации на основе образования гамет. Историческое возникновение пола медоносных пчёл относится к периоду эволюции цветковых растений, которые служили им пищей в виде нектара и пыльцы. Согласно данным науки об ископаемых – пчёлы на Земле существовали уже 42 миллиона лет назад. При расселении пчёл с Южной Азии к северу и востоку на новые территории, препятствием пчелам служили Гималайские горы, как непреодолимая преграда, а с юга был океан. В таких условиях единственной дорогой для расселения оставался западное направление в Европу. В тот период в Европе преобладал жаркий субтропический климат с богатой медоносной растительностью, лугов и лесов, благоприятной к размножению и расселению. Как отмечает И.А. Шабаршов (1990) начавшийся на Земле процесс горообразования. Например, как на Кавказе, вызвал похолодание на всей планете. В результате в половине Европы вплоть до Урала началось оледенение, и пчёлы снова стали, переселятся на юг. С наступлением потепления пчёлы стали мигрировать к северу, но уже в более холодный климат, чем было до оледенения. Такие изменения внешней среды послужили сменой систем определения пола. Нынешняя система определения пола по–видимому, для пчёл больше подходит, так как матки откладывают неоплодотворенные яйца для выращивания трутней в нужном количестве в более крупные ячейки сот гнезда и только для периода размножения (роения). Пол у пчел, как и у большинства организмов, определяется в момент оплодотворения, при котором половые хромосомы попадают в зиготу. Следовательно, это свойство организма, обеспечивающее функцию воспроизводства потомства и передачу ему свою генетическую информацию на основе образования гамет. Эти наследственные три системы заменяли друг друга в эволюционном процессе с развитием пчелиной семьи в зависимости от условий существования и питания. При исходной системе определения пола половыми хромосомами «XY» из одной половины яиц отложенных маткой развивались самки, а из другой самцы. При этой системе пчёлы жили в одиночку. Сами устраивали себе гнездо, клали яйца, собирали корм и выкармливали личинок медом и пыльцой.
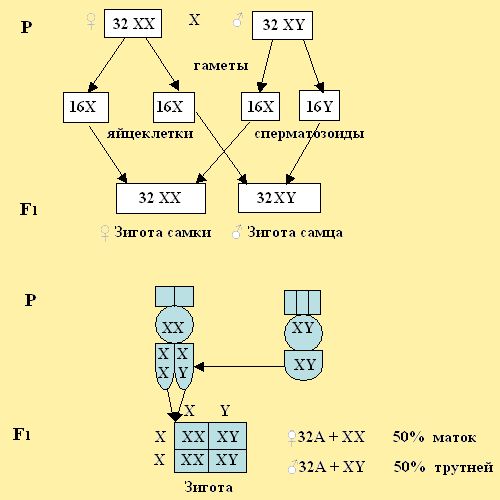
Вторая система возникла с прекращением одиночного образа жизни самок и образованием семьи с большим числом яйцекладущих самок и меньшим числом самцов от 50 до 2 – 5 – 8% (Шаскольский,1966). При второй системе у пчёл возник инстинкт не воспитывать трутней вне сезона по окончании периода размножения. На этой ступени эволюции дети, которые раньше покидали родительское гнездо, стали оставаться в гнезде, помогать матери строить ячейки, кормить личинок, а также сами откладывали яйца и добывали совместно пищу. В таких условиях стала создаваться специализация по видам выполнения работ. Одни матки клали яйца и реже вылетали за кормом, а другие переключались на добычу нектара и пыльцы для выкармливания расплода. Как отмечает Шабаршов (1990) то, что рабочие пчёлы не отличались от маток, подтверждается современной системой с появлением яйцекладущих пчел-трутовок, когда семья остается без матки. На этой стадии развития общественной жизни число плодных маток в семье превосходило число вспомогательных пчёл по добыче корма и воспитанию расплода. Многочисленное число яйцекладущих самок на этой ступени в семье жили дружно. Доказательством многоматочности служит закладка в семьях большого числа маточников в период размножения (роения) семьи. Более холодный климат северных районов заселяемых пчелами, для поддержания жизнедеятельности, потребовал иметь запас корма на период отсутствия в природе взятка. В связи с этим большая часть маток стали заготавливать больше корма, строить больше ячеек для яиц и отвлекаться от яйцекладки, а также пропускать брачные сроки своевременного спаривания с трутнями, что и послужило угасанием функции полового инстинкта. Семяприемник таких маток не получал спермы, уменьшался в объеме и как следствие превратился в рудиментарный. У яйцекладущих самок наоборот, обострялся инстинкт яйцекладки и угасал инстинкт по добыче корма уходу за расплодом, постройке сот и охрану гнезда. Специализация половых и других органов по сбору кормов, защиты гнезда постройке сот, по выращиванию большого количества расплода, послужила переходу семьи пчёл от многоматочного состояния к одноматочному. Во второй системе по окончании сезона взятка, а также при похолодании и резком обрыве взятка проявился инстинкт не воспитывать личинок и куколок трутней, а просто их выбрасывать из ячеек сот гнезда, не пускать в гнездо и выгонять из гнезда взрослых трутней. Третья система заменила вторую систему определения пола с развитием численности в семье рабочих особей, а также не выводить трутней вне сезонов роения (размножения). По существу, этот же инстинкт сохранился от второй системы не выводить трутней, а зародыши диплоидных трутней уничтожать (Шаскольский, 1966). Как определить пол пчелыПол наследуется как менделирующий признак (по законам Менделя). Сочетание половых хромосом в зиготе определяет пол будущего организма (рис. 150).
Рис. 150. Схема определения пола у человека. Половой хроматин у женщин. При созревании половых клеток в результате мейоза гаметы получают гаплоидный набор хромосом. В каждой яйцеклетке есть 22 аутосомы + Х-хромосома. Пол, который образуют гаметы, одинаковые по половой хромосоме, называют гомогаметным.
Такое определение пола характерно для человека и млекопитающих.
Рис. 151. Четыре типа определения пола.
Рис. 154. Основные клинические симптомы синдрома Тернера.
Рис. 155. Наследование признаков, сцепленных с полом, у дрозофилы.
Рис. 156. Родословные по признакам, сцепленным с половыми хромосомами.
Все девочки здоровы. Половина мальчиков больна гемофилией. Признаки, проявление которых различно у представителей разных полов, или проявляющиеся у одного пола, относятся к признакам, ограниченным полом. Эти признаки могут определяться генами, расположенными как в аутосомах, так и половых хромосомах. Возможность развития признака зависит от пола организма. Например, тембры голоса баритон и бас характерны только для мужчин. Проявление генов, ограниченных полом, связано с реализацией генотипа в условиях среды целостного организма. Помимо генов, ответственных за развитие вторичных половых признаков, которые в норме работают только у одного из полов, у другого они присутствуют, но «молчат». Функциональную активность целого ряда других генов определяет гормональная деятельность организма. Например, у быков есть гены, контролируйте продукцию молока и его качественные особенности (жирность, содержание белка и др.), но у быков они «молчат», а функционируют только у коров. Потенциальная способность быка давать высокомолочное потомство делает его ценным производителем молочного стада. Есть признаки, зависимые от пола. Гены, степень проявления которых определяется уровнем половых гормонов, называются генами, зависимыми от пола. Эти гены могут находиться не только в половых хромосомах, но и в любых аутосомах. Например, ген определяющий облысение, типичное для мужчин, локализован в аутосоме, и его проявление зависит от мужских половых гормонов. У мужчин этот ген действует как доминантный, а у женщин как рецессивный. Если у женщин этот ген в гетерозиготном состоянии, то признак не проявляется. Даже в гомозиготном состоянии у женщин этот признак слабее выражен, чем у мужчин. Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями. Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта. Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид. © 2018-2021 Все права на дизайн сайта принадлежат С.Є.А. Как определить пол пчелы
ТРУТНИ ТОЖЕ РАБОТАЮТ ДО СПАРИВАНИЯ.
Понятно. Но, если жизнь рабочей пчелы порядка 60 дней, то, извините, сколько времени работают трутни до своей половой зрелости? Неделю-две, что ли?!
ИХ МНОГО И ОНИ ЖИВУТ ДО ЗИМЫ,ПОТОМ ИХ ИЗ СЕМЬИ ИГОНЯЮТ.
А спариваются зимой, что ли? Или живут, растут, спариваются в начале лета, а потом просто кормятся, ничего не делая, до зимы? В то время, как рабочие девочки изводят себя на их кормёжке?! Немудрено, что зимой их из улья выгоняют. Надоели. Знакомое ощущение. Вполне поддерживаю!
Ой. и правда? По логике должны быть мужеского, но если по опыту и практике- женского.
ЖЕНСКОГО И С НЕДОРАЗВИТЫМИ РЕПРОДУКТИВНЫМИ ОРГАНАМИ. Как определить пол пчелы
В этом случае обычно винят матку (старая, больная, слабая) и стремятся ее заменить на другую, чаще со своей же пасеки, но все опять повторяется, поскольку истинная природа этого явления связана с особенностями генетических механизмов определения пола у пчелы медоносной. Впервые мысль о гапло-диплоидном механизме определения пола у пчелы была высказана Дзержоном более 150 лет тому назад (Dzierzon, 1861): матка и рабочие пчелы развиваются из оплодотворенных, а трутни — из неоплодотворенных яиц. Предположение Маккензена было блестяще экспериментально подтверждено работами Войке (1963) с помощью искусственного осеменения пчелиных маток трутнями-братьями. Эти результаты были доложены на XIX Международном конгрессе по пчеловодству. Оригинальность данной работы заключается в том, что автор определял пол насекомого еще в запечатанном расплоде по анатомическим особенностям зачатков половых органов личинок. Изучался расплод только тех маток, в семьях которых отмечалось около 50% пестрого расплода. Было обнаружено, что половина зародышей потомства таких маток несет признаки женского пола, а другая половина — мужского. Таким путем удалось доказать, что эмбриональная гибель зародыша связана с геном пола. Автором было также высказано предположение о том, что диплоидные зародыши трутней погибают не сами по себе: их распознают и уничтожают рабочие пчелы сразу после выхода из яиц. Почти одновременно с этой работой Лейдлов с соавторами (Laidlow et al., 1956), изучая распространение полулетальных генов в бразильской популяции более 60 пчелиных семей, обнаружили 12 аллелей этого гена. Эволюция генетических механизмов определения пола у пчел. Таким образом у пчелы медоносной описано сосуществование двух систем определения пола: гапло-диплоидной и множественных аллелей гена пола. Предполагается, что это обстоятельство отражает эволюционные преобразования генетической системы определения пола, связанные с переходом пчелы от одиночного к семейному существованию (Шаскольский, 1971). Согласно автору, можно предположить существование трех этапов в становлении (совершенствовании) генетических механизмов систем определения пола у пчелы. Эти системы эволюционировали в связи с развитием и усложнением пчелиной семьи. I этап. Исходная система — это самый древний механизм становления пола, определяется гетерохромосами (ХУ или ХО). Он широко распространен в мире насекомых. Такая система обеспечивает равновероятное появление особей мужского и женского пола в соотношении 1:1 при диплоидности самцов и самок. Замена первой системы на вторую связана с прекращением одиночного существования и образованием семьи, специализацией особей в ней (матка, рабочие пчелы и трутни), резким сокращением числа размножающихся самок, уменьшение потребности в самцах сопровождалось совершенствованием генетических механизмов определения пола. III этап. Гапло-диплоидная система определения пола. Появление этой системы связано, по-видимому, с дальнейшим развитием семьи, увеличением численности ее особей до нескольких тысяч, при этом потребность в трутнях ограничивается только сезонным размножением. Инстинкт удаления ненужных трутней сохраняется и в другом вариенте: когда рабочие особи при резком похолодании или сокращении взятка выбрасывают уже зрелый трутневый расплод. Гапло-диплоидный механизм регуляции пола позволяет пчелиной семье довольно легко регулировать время вывода трутней, а также их численность. Однако наряду с этим механизмом механизм множественных аллелей гена пола сохраняется в виде рудиментарной системы, которая заявляет о себе при близкородственном разведении феноменом пестрого расплода. Геном трутня как генетический полигон. Переход на гапло-диплоидный механизм определения пола дал огромные преимущества медоносной пчеле в смысле цитогенетического механизма адаптации и ознаменовал исключительную роль трутней в этом процессе. Гаплодная генетическая система самцов, обладающая одинарным набором хромосом, а следовательно и генов, представляет собой полигон, на котором выявляются все вредные рецессивные гены, снижающие жизнеспособность и адаптационный потенциал вида. Их проявление приводит к гибели особей, носителей этих генов. Таким образом, популяция имеет возможность освободиться от так называемого «генетического груза». Именно науровне генетической системы трутней идет отбор наиболее приспособленных к конкретным условиям существования генетических комбинаций. Поэтому, уничтожая трутневый расплод, мы тем самым препятствуем реализации одной из самых совершенных программ адаптации вида, которая совершенствовалась миллионы лет эволюции. Именно она придает генетическому аппарату пчелы удивительную пластичность, которая позволила Apis mellifera освоить все континенты, за исключением Антарктиды. Интересно отметить, что гапло-диплоидный механизм определения пола широко распространен среди насекомых (более чем у 200 тыс. видов). Рассмотрим на примере возможность комбинирования половых аллелей в локальной популяции.
Очевидно, чтобы избежать высокой концентрации одноименных аллелей на пасеке при искусственном выводе маток, необходимо использовать несколько донорских семей. Это особенно важно на изолированных пасеках. В большинстве случаев пчелы сами преодолевают эту проблему. Случные пункты нередко бывают удалены от пасеки на значительные расстояния, что уменьшает вероятность родственного спаривания. С другой стороны, полиандрия (осеменение несколькими трутнями), несомненно, ценное биологическое приспособление, уменьшающее вероятность концентрации одноименных аллелей в одной популяции. Проблема идентификации половых аллелей. Идентификация половых аллелей приобретает большое значение при искусственном осеменении маток. В этом случае генетический контроль производителей — важное условие селекционно-генетических исследований. Секция «Биотехнологические Особенности генетики пчелГенетика — раздел биологической науки, посвященный изучению наследственности и изменчивости организмов. Основные закономерности наследственности и изменчивости изучает общая генетика, а специфические, присущие отдельным видам, — частная генетика. Поскольку частная генетика медоносной пчелы изучена еще недостаточно, то в настоящей главе имеющиеся по этому вопросу данные приводятся в виде дополнений к информации по основным вопросам общей генетики. Вопросы общей генетики. Под наследственностью имеется в виду материальная и функциональная преемственность между поколениями организмов или способность воспроизводства потомками признаков и свойств своих родителей. Осуществляемая при этом передача наследственной информации от поколения к поколению называется наследуемостью. Появление различий между родителями и потомками в тех или иных признаках или свойствах называется изменчивостью, которая бывает наследственной (генотипической) и ненаслсдственной (модификационной). Общая изменчивость организма, представляющая собой совместное проявление как генотипической, так и модификационной изменчивости, называется фенотипической. Под генотипом понимают совокупность наследственных задатков организма, а под фенотипом — их внешнее проявление в конкретных условиях. Основными носителями наследственной информации являются хромосомы — специфические элементы клеточного ядра, состоящие из гигантских молекул дезоксирибонуклеиновой кислоты (ДНК) и белков. Молекула ДНК представляет собой две полинуклеотидные цепи, образующие спираль. В состав нуклеотидов, образующих ДНК, входят азотистые основания, остаток фосфорной кислоты и дезоксирибоза. В процессе передачи наследственной информации важную роль играют также молекулы рибонуклеиновой кислоты (РНК). Элементарной единицей наследственной информации является ген, представляющий собой участок хромосомы или лежащей в ее основе молекулы ДНК. Гены контролируют с помощью ферментов синтез как белковых, так и небелковых веществ, от которых зависит формирование всех признаков организма. Ген, контролирующий тот или иной признак, занимает вполне определенное место в соответствующей хромосоме, которое называется локусом. Каждый ген имеет не менее двух форм состояния, называемых аллелями: один доминантный (подавляющий влияние альтернативного аллеля); второй рецессивный (влияние которого полностью или частично подавляется доминантным аллелем). В результате мутаций какого-либо гена может возникнуть несколько его аллелей, т. е. целая серия, по-разному влияющих на соответствующий признак. Каждый новый аллель, в свою очередь, также может мутировать и дать начало новому аллелю. Образовывать серии могут как доминантные, так и рецессивные аллели. Аллели из множественных серий рассредоточены между разными особями одной и той же популяции, а у конкретного диплоидного организма в хромосомах может присутствовать только одна пара аллелей из данной серии. Действие множественных аллелей также подчиняется законам Менделя. Чем больше аллелей в данной серии, тем больше вариантов попарного комбинирования между ними. Наряду с генами, имеющими два и более аллелей, существуют и неаллельные гены, к которым относятся так называемые гены-модификаторы. Сами по себе они никакого влияния на данный признак не оказывают, но регулируют воздействие главного гена, ослабляя или усиливая его. Главными генами, или олигогенами, называют те из них, которые предопределяют формирование качественных, т. е. альтернативных, признаков. Условия внешней среды слабо влияют на силу проявления олигогена. Иногда олигоген может одновременно влиять и на определенный качественный, и на определенный количественный признак. Способность одного гена влиять сразу на несколько признаков называется плейотропией. Плейотропия свойственна многим генам и лежит в основе генотипической корреляционной изменчивости. Полигены (полимерные гены) относятся к категории неаллельных и контролируют проявление количественных признаков. Важнейшая особенность полигенов заключается в аддитивном (суммарном) воздействии их на этот признак. Результат воздействия полимерных генов на определенный признак организма называется аддитивным эффектом. Для качественных признаков характерна прерывистая изменчивость, для количественных — непрерывная. Условия окружающей среды вызывают сильную модификационную изменчивость количественных признаков. У количественных (полигенных) признаков ни доминирования, ни эпистаза (взаимодействие генов, когда аллель одного гена подавляет влияние аллелей другого, регулирующего совершенно иной признак) быть не может. Генетическая (наследственная) изменчивость подразделяется на мутационную и комбинационную. Мутационная изменчивость вызывается структурными преобразованиями генов или хромосом, в результате которых появляются новые признаки и свойства. Мутация может произойти спонтанно или явиться результатом воздействия тех или иных физических или химических факторов (мутагенов). Комбинационная изменчивость наблюдается при скрещивании разных типов и пород животных, когда образуются новые сочетания родительских генов (новый генотип). Новые комбинации аллелей этих генов возникают в процессе кроссинговера (взаимного обмена одинаковыми участками между гомологичными хромосомами), обычно происходящего во время мейоза (редукционного деления клеточных ядер). Комплекс хромосом, типичный для соматических клеток данного вида животного организма по количеству, размерам и особенностям строения, называют кариотипом. Такой комплекс, или набор хромосом, именуют диплоидным, или двойным, в отличие от гаплоидного, или одинарного, набора, типичного для половых клеток (гамет). Разные виды организмов имеют только им присущее количество хромосом в кариотипе. В частности, у маток и рабочих особей медоносной пчелы (Apis mellifera L.) и пчелы средней индийской (Apis cerana indica F.) диплоидный набор содержит 32 хромосомы, гаплоидный — 16. У трутней этих двух видов и в соматических, и в половых клетках содержится по 16 хромосом. У пчелы гигантской (Apis dorsata) и пчелы карликовой (Apis florea) — представителей этого же рода количество хромосом в 2 раза меньше, соответственно у самок 16 и 16, у самцов — 8 и 8. Кариотип каждого вида неизменен (за исключением наблюдаемой полиплоидизации) и потому довольно часто используется для установления филогенетического родства. Известно, что в хромосомах содержится вся наследственная информация, однако в диплоидной клетке каждый ген представлен обоими аллелями, а в гаплоидной — одним. Изменение кариотипа может вызвать только геномная или хромосомная мутация, в результате которой возникают особи-полиплоиды. Механизм полиплоидизации можно представить себе на иногда наблюдаемых примерах, когда в результате митоза (деление клеток в процессе роста организма) число хромосом удвоилось, но разделения ядра не последовало — клетка превратилась в полиплоидную. Некоторые авторы отрицают наличие мейоза у трутней, а другие полагают, что в данном случае этот процесс видоизменил свой характер, поскольку из одного сперматогония образуется один спермий. У пчел, муравьев и ряда других представителей отряда перепончатокрылых половые хромосомы отсутствуют, а самцы развиваются партеногенетически. У этих насекомых имеет место гаплоидное определение пола, т. е. самки развиваются из оплодотворенных яиц и становятся диплоидными, а гаплоидные самцы — из неоплодотворенных. Развитие начинается с дробления зиготы, которое в первом случае стимулируется оплодотворением, а во втором — какими-то еще недостаточно выявленными внутренними или внешними активаторами партеногенетического развития. Поэтому у трутней нет ни отцов, ни сыновей, однако есть дедушки, матери, дочери и внуки. Взаимодействие женского генотипа с факторами внешней среды (характер личиночного кормления, размер и форма ячейки, температурный режим) предопределяет формирование двух различных каст: или полноценной самки (матки пчелиной), или рабочей особи. Дзержон еще в 1845 г. открыл явление мужского партеногенеза у медоносной пчелы. Однако в последние десятилетия появились и новые гипотезы, дополняющие наши представления о половой детерминации у медоносной пчелы. Уайтинг (1943) с помощью тесного инбридинга получил диплоидных самцов у паразитической осы харбобракон. Самцы у этого вида, как и у всех перепончатокрылых, развиваются партеногенетически, т. е. из неоплодотворенных гаплоидных яиц. Позже Маккензен (1951, 1955) опубликовал результаты своих работ по изучению близкородственного разведения у медоносных пчел и показал, что при этом наблюдался примерно тот же процент нежизнеспособных яиц, что и в аналогичном случае у осы харбобракон. Однако диплоидных трутней тогда обнаружить еще никому не удалось. В 1957 г. Ротенбулер нашел трутней, у которых имела место комбинация диплоидных и гаплоидных тканей. Однако первым, кто сумел получить диплоидных трутней, был Войне (1965). В результате ряда исследований и наблюдений, проведенных этими авторами, была создана следующая гипотеза. Половых хромосом у медоносной пчелы нет, однако найден генетический локус, обозначенный как χ-локус, который обеспечивал половую детерминацию у перепончатокрылых задолго до появления мужского партеногенеза. Этот ген представлен серией множественных аллелей, количество которых, по данным различных авторов, колеблется в пределах от 12 до 25. Новый способ детерминации пола (развитие женских особей из оплодотворенных яиц и мужских — из неоплодотворенных) возник очень давно, но и предшествовавший ему механизм (серия множественных аллелей полового гена) полностью не исчез и в некоторых случаях проявляет себя довольно ощутимо. Самки (матки и рабочие особи), будучи диплоидными, всегда имеют два половых аллеля и гетерозиготны по этому гену, т. е. содержат два разных его аллеля. Такая матка, спарившись с гаплоидным трутнем, который имеет только один половой аллель, будет продуцировать два типа женских особей и два типа мужских (гаплоидных). Если это потомство спарится между собой во всех возможных сочетаниях, то половина маток. Вторая половина таких же маток, но спарившихся с трутнями, половые аллели которых аналогичны одному из аллелей матки-партнера, будет продуцировать таких же гаплоидных трутней, как и в предыдущем случае, а также два типа диплоидных особей: 50% гетерозиготных и 50% гомозиготных. Все гетерозиготные по гену пола особи станут самками (самки всегда гетерозиготны по гену пола), а гомозиготные — диплоидными самцами. Личинки бипарентальных диплоидных трутней будут съедены пчелами-кормилицами через несколько часов после выхода их из яиц. Войке считает, что причина поедания личинок диплоидных трутней заключается в том, что они выделяют гормон каннибализма. Ему удалось вырастить диплоидных трутней из этих личинок (своевременно отобранных у пчел) в лабораторных условиях (в термостате), однако они отличались низкой жизнеспособностью. Матки-матери диплоидных трутней откладывали в трутневые ячейки неоплодотворенные яйца, из которых выводились обычные гаплоидные трутни. Диплоидные трутни крупнее гаплоидных, во время созревания их спермиев не происходит редукции хромосом, поэтому они формируются также диплоидными. Рассмотрим, как эта биологическая особенность медоносной пчелы выглядит в конкретных условиях практического пчеловодства (по Д. В. Шаскольскому). Пчелиная матка, как известно, диплоид, так как получает по 16 хромосом (по гаплоидному набору) и от отца и от матери. В откладываемое ею яйцо попадает либо отцовский, либо материнский комплект хромосом. Поэтому одна половина откладываемых ею яиц получает материнский аллель, а вторая — отцовский. У разных маток и трутней на пасеке, как правило, наблюдается много разных аллелей пола. Поэтому в большинстве случаев яйцо с одним аллелем оплодотворяется спермием, содержащим какой-то другой аллель гена пола. В результате возникнет гетерозигота, из которой разовьется нормальная самка. Вместе с тем в некоторых случаях яйцо будет оплодотворено спермием, несущим точно такой же аллель гена пола, т. е. оно станет гомозиготным по этому аллелю. Вышедшая из этого яйца диплоидная трутневая личинка вскоре будет съедена пчелами. Допустим, матка спарилась всего с восемью трутнями, из которых три имели такие же аллели пола, как и один из двух имеющихся у матки, а пять — совершенно иные. В этом случае большинство оплодотворенных яиц будут гетерозиготными по гену пола, из которых выведутся нормальные женские особи, а меньшинство, т. е. гомозиготные, будут съедены. Количество таких яиц при 16 возможных вариантах сочетания из двух упоминавшихся выше материнских и 8 отцовских половых аллелей теоретически равняется 3/16, или 19%. Этот показатель может несколько варьировать из-за неодинакового количества спермиев от разных трутней, попавших в сперматеку матки, или из-за неравномерного их перемешивания в сперматеке. Как известно, матка спаривается с 10—15 трутнями. В качестве крайне редких вариантов можно ожидать, что очень немногие матки либо не будут откладывать гомозиготных диплоидных яиц (если они спарятся с трутнями, имеющими аллели, несходные с их собственными), либо будут репродуцировать их в максимальном количестве (до 50%, если их все трутни-партнеры будут обладать точно такими же аллелями), а большинство маток по проценту откладки таких яиц будет находиться где-то между этими крайними значениями. Медопродуктивность таких пчелиных семей будет снижаться пропорционально количеству откладываемых диплоидных яиц, гомозиготных по аллелю полового гена. Чтобы практически определить долю таких яиц, откладываемых матками, на пасеке выборочно выделяют пятую или даже десятую часть пчелиных семей. В эти семьи помещают по рамочному изолятору из разделительной решетки. В такой изолятор помещают матку и рамку с хорошим сотом для откладки яиц. Через каждые 3-е суток сот изымают из изолятора, переставляя в гнездо, а вместо него ставят новый. Когда пчелы запечатают расплод на этих сотах, то подсчитывают количество запечатанных ячеек и находящихся между ними пустых (где находились съеденные пчелами гомозиготные диплоидные трутневые личинки). Можно облегчить эту работу и повысить точность подсчета, если соты сфотографировать, а затем произвести подсчет по фотографиям. После подсчета определяют процент и тех, и других ячеек, т. е. процент откладываемых маткой гетеро — и гомозиготных по аллелям пола яиц. Если же матку и соты в изолятор не помещать, то она вместо съеденных пчелами гомозиготных личинок отложит сразу же другие яйца, и сделать точные определения не удастся. Можно применить и упрощенный способ, суть которого заключается в том, что в проверяемую семью подставляют хороший сот. Через неделю по возрасту личинок определяют (условно) дату откладки яиц в его ячейки, через 12 дней после этой даты сот фотографируют. Расплод, развившийся из гетерозиготных по аллелю пола яиц, в это время будет уже запечатанным, а развившийся из яиц, отложенных вместо съеденных гомозиготных личинок, — еще нет. Как вытекает из вышеизложенного, идентичные половые аллели, прежде всего, встречаются у близкородственных маток и трутней и гомозиготные диплоидные яйца возникают именно при их спаривании (брат Х сестра). У инбредных линий, например, количество половых аллелей сокращается всего лишь до 2. Поэтому спаривание маток и трутней в воздухе и на значительном удалении от пасеки, а также тот факт, что матка спаривается не с одним, а с несколькими трутнями, следует рассматривать, как возникшее в процессе эволюции приспособление против перехода половых аллелей в летальное гомозиготное состояние. Чтобы не допустить сколь-нибудь заметного ущерба, возникающего при этом, необходимо принимать специальные меры: Есть и другие гипотезы относительно генетического механизма половой дифференциации у медоносной пчелы. Так, Керр и Нельсон (1967) считают, что этот процесс контролируют два набора генов, один из которых действует в самом начале эмбрионального развития особей, а второй — в предкуколочной стадии (во взаимодействии с ювенильным гормоном). Кунхе и Керр (1957) предполагают существование гена «самковостн» (f). При ff возникает кумулятивный эффект lF. У гена «самцовости» не бывает кумулятивного эффекта (m = lM). По мнению Гонсалвеса, характер поведения пчел контролируют 6 генов. Есть предпосылки полагать, что у пчел миролюбие доминирует над агрессивностью, а низкий кубитальный индекс — над высоким (Ритир). Как известно, размер ячеек естественных построек является породным признаком (Альбер), а большой размер ячейки доминирует над меньшим. Признак высоких санитарных способностей (очистки ячеек и гнезда от больных и погибших личинок и куколок) по отношению к признаку отсутствия этого качества является рецессивным (Ротенбулср). Способность активно собирать пыльцу с люцерны наследуется промежуточно и контролируется несколькими генами. Аллель отсутствия опушения тела пчелы является рецессивным по отношению к аллелю дикого типа нормального опушений. Подавляющее большинство авторов приводят убедительные доказательства о том, что желтизна в окраске тела пчелы доминирует над черным или коричневым цветом, однако изредка приводятся и данные противоположного характера. В настоящее время разработана техника оценки множественного аллелизма, заключающаяся в том, что мутантная матка с неизвестным генотипом осеменяется спермой мутантных трутней с известным генотипом. В результате использования этой методики сокращается время и число необходимых скрещиваний. Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.  В жизни всегда есть моменты, когда нам приходится принимать  В учебной среде, где знания и информация становятся |



















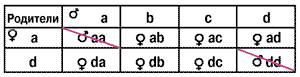 Все гетерозиготные по гену пола особи будут нормальными особями женского пола, а гомозитогные — диплоидными трутнями, которых пчелы уничтожат, что и приведет к картине пестрого расплода. Если трутни окажутся братьями матки, несущими одинаковые с ней половые аллели, то процент пестрого расплода будет максимальным — 50%.
Все гетерозиготные по гену пола особи будут нормальными особями женского пола, а гомозитогные — диплоидными трутнями, которых пчелы уничтожат, что и приведет к картине пестрого расплода. Если трутни окажутся братьями матки, несущими одинаковые с ней половые аллели, то процент пестрого расплода будет максимальным — 50%.
