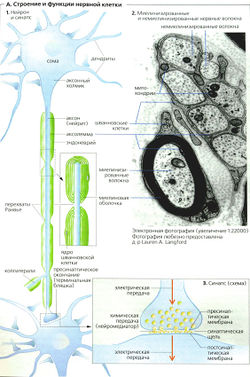
2.1. Нейроны
Нейроны (рис. 2) являются возбудимыми клетками нервной системы. В отличие от глиальных клеток они способны возбуждаться (генерировать потенциалы действия) и проводить возбуждение. Нейроны высокоспециализированные клетки и в течение жизни не делятся.
В нейроне выделяют тело (сому) и отростки. Сома нейрона имеет ядро и клеточные органоиды (рис. 2, 1). Основной функцией сомы является осуществление метаболизма клетки.
1 — сома (тело) нейрона; 2 — дендрит; 3 — тело Швановской клетки; 4 — миелинизированный аксон; 5 — коллатераль аксона; 6 — терминаль аксона; 7 — аксонный холмик; 8 — синапсы на теле нейрона
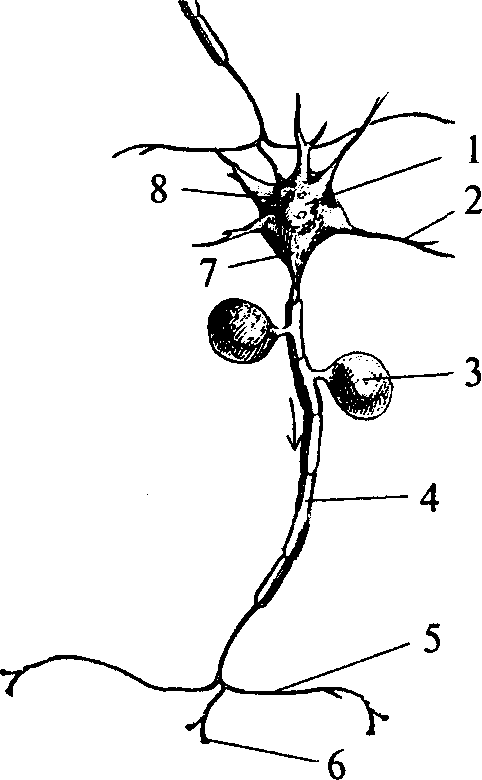
Число отростков у нейронов различно, но по строению и выполняемой функции их делят на два типа. Одни — короткие, сильно ветвящиеся отростки, которые называются дендритами (от dendro — дерево, ветвь). Нервная клетка несет на себе от одного до множества дендритов (рис. 2, 2). Основной функцией дендритов является сбор информации от множества других нейронов. Ребенок рождается с ограниченным числом дендритов (межнейронных связей), и увеличение массы мозга, которое происходит на этапах постнатального развития, реализуется за счет увеличения массы дендритов и глиальных элементов.
Другим типом отростков нервных клеток являются аксоны (рис. 2, 4). Аксон в нейроне один и представляет собой более или менее длинный отросток, ветвящийся только на дальнем от сомы конце. Эти ветвления аксона называются аксонными терминалами (окончаниями) (рис. 2, 6). Место нейрона, от которого начинается аксон (рис. 2, 7; 6, 2), имеет особое функциональное значение и называется аксонным холмиком (рис. 2, 7). Здесь генерируется потенциал действия — специфический электрический ответ возбудившейся нервной клетки. Функцией же аксона является проведение нервного импульса к аксонным терминалям. По ходу аксона могут образовываться его ответвления — коллатерали (рис.2, 5). В месте отхождения коллатерали (бифуркации) импульс «дублируется» и распространяется как по основному ходу аксона, так и по коллатерали.
Часть аксонов центральной нервной системы покрывается специальным электроизолирующим веществом — миелином. Миелинизацию аксонов осуществляют клетки глии. В центральной нервной системе эту роль выполняют олигодендроциты, в периферической — Шванновские клетки (рис. 2, 3; 3, 2), являющиеся
1 — связь между телом клетки глии и миелиновой оболочкой; 2 — олигодендроцит; 3 — гребешок; 4 — плазматическая мембрана; 5 — цитоплазма олигодендроцита; 6 — аксон нейрона; 7 — перехват Ранвье; 8 — мезаксон; 9 — петля плазматической мембраны
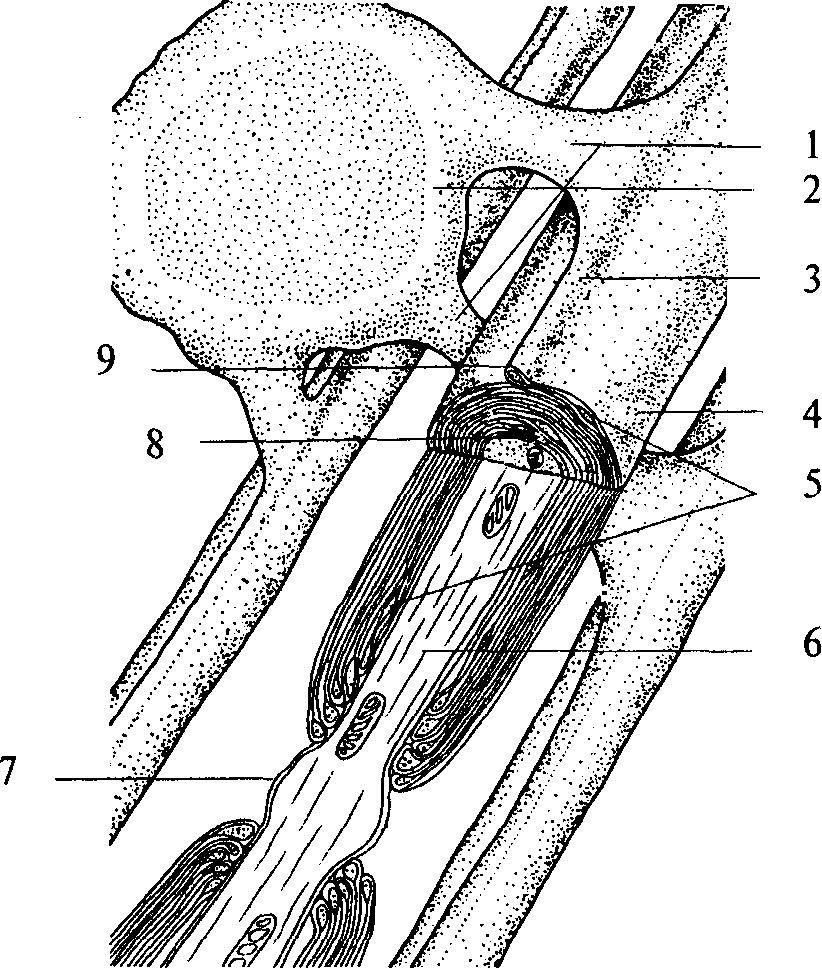
разновидностью олигодендроцитов. Олигодендроцит оборачивается вокруг аксона, образуя многослойную оболочку. Миелинизации не подвергается область аксонного холмика (рис. 2, 7) и терминали аксона. Цитоплазма глиальной клетки выдавливается из межмембранного пространства в процессе «обертывания». Таким образом, миелиновая оболочка аксона состоит из плотно упакованных, перемежающихся липидных и белковых мембранных слоев (рис. 3, 4). Аксон не сплошь покрыт миелином. В миелиновой оболочке существуют регулярные перерывы — перехваты Ранвье (рис. 3, 7). Ширина такого перехвата от 0,5 до 2, 5 мкм. Функция перехватов Ранвье — быстрое скачкообразное (сальтаторное) распространение потенциалов действия, осуществляющееся без затухания. В центральной нервной системе аксоны различных нейронов, направляющиеся к одной структуре, образуют упорядоченные пучки —
1 — аксон пресинаптического нейрона; 2 — микротрубочки; 3 — синаптический пузырек (везикула); 4 — синаптическая щель; 5 — дендрит постсинаптического нейрона; 6 — рецептор для медиатора; 7 — постсинаптическая мембрана; 8 — пресинаптическая мембрана; 9 — митохондрия
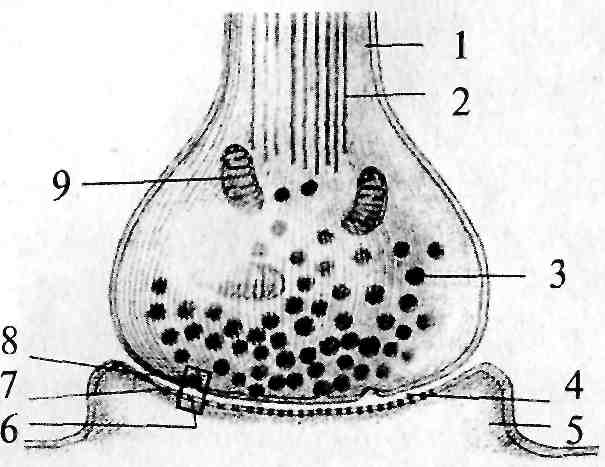
проводящие пути. В подобном проводящем пучке аксоны направляются «параллельным курсом» и часто одна глиальная клетка образует оболочку нескольких аксонов. Поскольку миелин является веществом белого цвета, то проводящие пути нервной системы, состоящие из плотно лежащих миелинизированных аксонов, образуют белое вещество мозга. В сером же веществе мозга локализуются тела клеток, дендриты и немиелинизированные части аксонов.
В пределах центральной нервной системы каждая терминаль аксона оканчивается на дендрите, теле или аксоне других нейронов. Контакты между клетками подразделяются в зависимости от того, чем они образованы. Контакт, образуемый аксоном на дендрите, называется аксо-дендритным; аксоном на теле клетки — аксо-соматическим; если он образован двумя аксонами, то называется аксо-аксональным, а двумя дендритами — дендро-дендритным.
За пределами ЦНС терминали могут заканчиваться как на нейронных элементах, так и на других возбудимых клетках (мышечных или железистых). В любом случае между нейроном и последующей клеткой образуется специфический контакт — синапс (рис. 4, 4). В образовании синапса участвуют как аксонная терминаль (пресинаптическая часть), так и мембрана последующей клетки (постсинаптическая часть). Синапс состоит из пресинаптической бляшки (расширение терминали аксона), оканчивающейся пресинаптической мембраной (рис. 4, 8), и постсинаптической мембраны (участка мембраны постсинаптической клетки, лежащего под синаптической бляшкой) (рис. 4, 7). Между пресинаптической и постсинаптической мембранами расположена синаптическая щель (рис. 4, 4).
От ее величины зависит тип передачи информации через синапс. Если расстояние между мембранами нейронов не превышает
2—4 нм или они контактируют между собой, то такой синапс является электрическим, поскольку подобное соединение обеспечивает низкоомную электрическую связь между этими клетками, позволяющую электрическому потенциалу непосредственно или электротонически передаваться от клетки к клетке. Доля электрических синапсов в ЦНС позвоночных очень мала.
Чаще всего мембраны нейронов расположены в непосредственной близости друг к другу и разделены обычным межклеточным пространством (щелью шириной примерно 20 нм) — смежное соединение. Такая смежность мембран облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов. Эти соединения нейронов относят к химическим синапсам.
В пресинаптическом окончании химического синапса находятся пузырьки — везикулы (рис. 4, 5), содержащие вещество — передатчик, называемое медиатором. В момент прихода к синаптической бляшке электрического импульса везикулы открываются в пресинаптическую щель, выбрасывая туда медиатор. Медиатор диффундирует через щель и на постсинаптической мембране взаимодействует с рецептором (рис. 4, 6), специфически чувствительным к медиатору, при этом возникает постсинаптический потенциал. Исключением из данного правила являются пептидергические нейроны, не имеющие в пресинаптической области везикул, так как медиатор-пептид синтезируется в соме нейрона и транспортируется по аксону в зону контакта.
Таким образом, информация в нервной системе передается только в одном направлении (от пресинаптического нейрона к постсинаптическому) и в этом процессе участвует биологически активное вещество — медиатор.
До 50-х годов XX столетия к медиаторам относили две группы низкомолекулярных соединений: амины (ацетилхолин, адреналин, норадреналин, серотонин, дофамин) и аминокислоты (гамма-аминомасляная кислота, глутамат, аспартат, глицин). Позже было показано, что специфическую группу медиаторов составляют нейропептиды, которые могут выступать также и в качестве нейромодуляторов (веществ, изменяющих величину ответа нейрона на стимул).
В настоящее время известно, что нейрон может синтезировать и выделять несколько нейромедиаторов (сосуществующие медиаторы). Такое представление о химическом кодировании вошло в основу принципа множественности химических синапсов. Нейроны обладают нейромедиаторной пластичностью, т.е. способны менять основной медиатор в процессе развития. Сочетание медиаторов может быть неодинаковым для разных синапсов.
В нервной системе существуют особые нервные клетки — нейросекреторные. Они имеют типичную структурную и функциональную (т.е. способность проводить нервный импульс) нейрональную организацию, а их специфической особенностью является нейросекреторная функция, связанная с секрецией биологически активных веществ. Функциональное значение этого механизма состоит в обеспечении регуляторной химической коммуникации между центральной нервной и эндокринной системами, осуществляемой с помощью нейросекретируемых продуктов.
В процессе эволюции клетки, входящие в состав примитивной нервной системы, специализировались в двух направлениях: обеспечение быстро протекающих процессов, т.е. межнейронное взаимодействие, и обеспечение медленно текущих процессов, связанных с продукцией нейрогормонов, действующих на клетки-мишени на расстоянии. В процессе эволюции из клеток, совмещающих сенсорную, проводниковую и секреторную функции, сформировались специализированные нейроны, в том числе и нейросекреторные. Следовательно, нейросекреторные клетки произошли не от нейрона как такового, а от их общего предшественника — пронейроцита беспозвоночных животных. Эволюция нейросекреторных клеток привела к формированию у них, как и у классических нейронов, способности к процессам синаптического возбуждения и торможения, генерации потенциала действия.
Для млекопитающих характерны мультиполярные нейросекреторные клетки нейронного типа, имеющие до 5 отростков. Такого типа клетки имеются у всех позвоночных, причем они в основном составляют нейросекреторные центры. Между соседними нейросекреторными клетками обнаружены электротонические щелевые контакты, которые, вероятно, обеспечивают синхронизацию работы одинаковых групп клеток в пределах центра.
Аксоны нейросекреторных клеток характеризуются многочисленными расширениями, которые возникают в связи с временным накоплением нейросекрета. Крупные и гигантские расширения называются «телами Геринга». В пределах мозга аксоны нейросекреторных клеток, как правило, лишены миелиновой оболочки. Аксоны нейросекреторных клеток обеспечивают контакты в пределах нейросекреторных областей и связаны с различными отделами головного и спинного мозга.
Одна из основных функций нейросекреторных клеток — это синтез белков и полипептидов и их дальнейшая секреция. В связи с этим в клетках подобного типа чрезвычайно развит белоксинтезирующий аппарат — это гранулярный эндоплазматический ретикулум и аппарат Гольджи. Сильно развит в нейросекреторных клетках и лизосомальный аппарат, особенно в периоды их интенсивной деятельности. Но самым существенным признаком активной деятельности нейросекреторной клетки является количество элементарных нейросекреторных гранул, видимых в электронном микроскопе.
Коллатерали аксона что это
Функциональная единица нервной системы — нервная клетка, нейрон. Нейроны способны генерировать электрические импульсы и передавать их в виде нервных импульсов. Нейроны образуют между собой химические связи — синапсы. Соединительная ткань нервной системы представлена нейроглией (дословно— «нервная глия»). Клетки нейроглии так же многочисленны, как и нейроны, и выполняют трофическую и опорную функции.
Миллиарды нейронов формируют поверхностный слой — кору— полушарий головного мозга и полушарий мозжечка. Кроме того, в толще белого вещества нейроны образуют скопления—ядра.
Практически все нейроны ЦНС мультиполярны: сома (тело) нейронов характеризуется наличием нескольких полюсов (вершин). От каждого полюса, за исключением одного, отходят отростки — дендриты, которые образуют многочисленные разветвления. Дендритные стволы могут быть гладкими или образовывать многочисленные шипики. Дендриты образуют синапсы с другими нейронами в области шипиков или ствола дендритного дерева.
От оставшегося полюса сомы отходит отросток, проводящий нервные импульсы,— аксон. Большинство аксонов формирует коллатеральные ветви. Концевые ветви образуют синапсы с нейронами-мишенями.
Нейроны образуют два основных типа синаптических контактов: аксодендритические и аксосоматические. Аксодендритические синапсы в большинстве случаев передают возбуждающие импульсы, а аксосоматические — тормозящие.
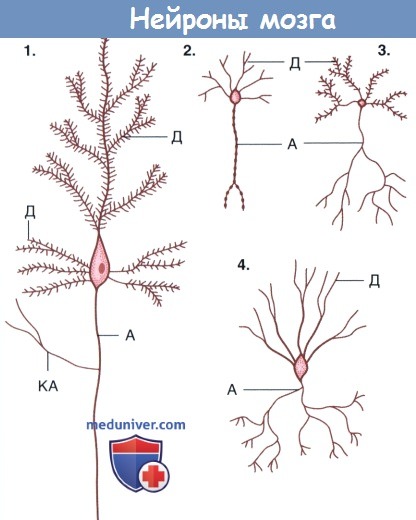 Формы нейронов мозга.
Формы нейронов мозга.
(1) Пирамидальные нейроны коры полушарий.
(2) Нейроэндокринные нейроны гипоталамуса.
(3) Шипиковые нейроны полосатого тела.
(4) Корзинчатые нейроны мозжечка. Дендриты нейронов 1 и 3 образуют шипики.
А — аксон; Д — дендрит; КА — коллатерали аксона. 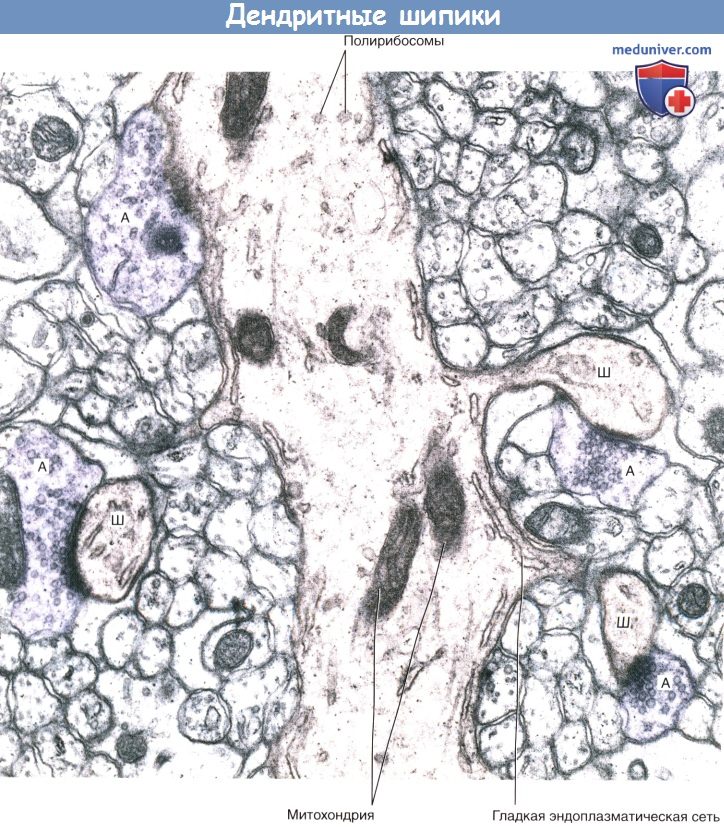 Дендритные шипики.
Дендритные шипики.
Срез мозжечка, на котором имеются дендриты гигантских клеток Пуркинье, образующие шипики.
В поле зрения различимы три шипика (Ш), образующие синаптические контакты с булавовидными расширениями аксонов (А).
Четвертый аксон (слева вверху) образует синапс с дендритным стволом. 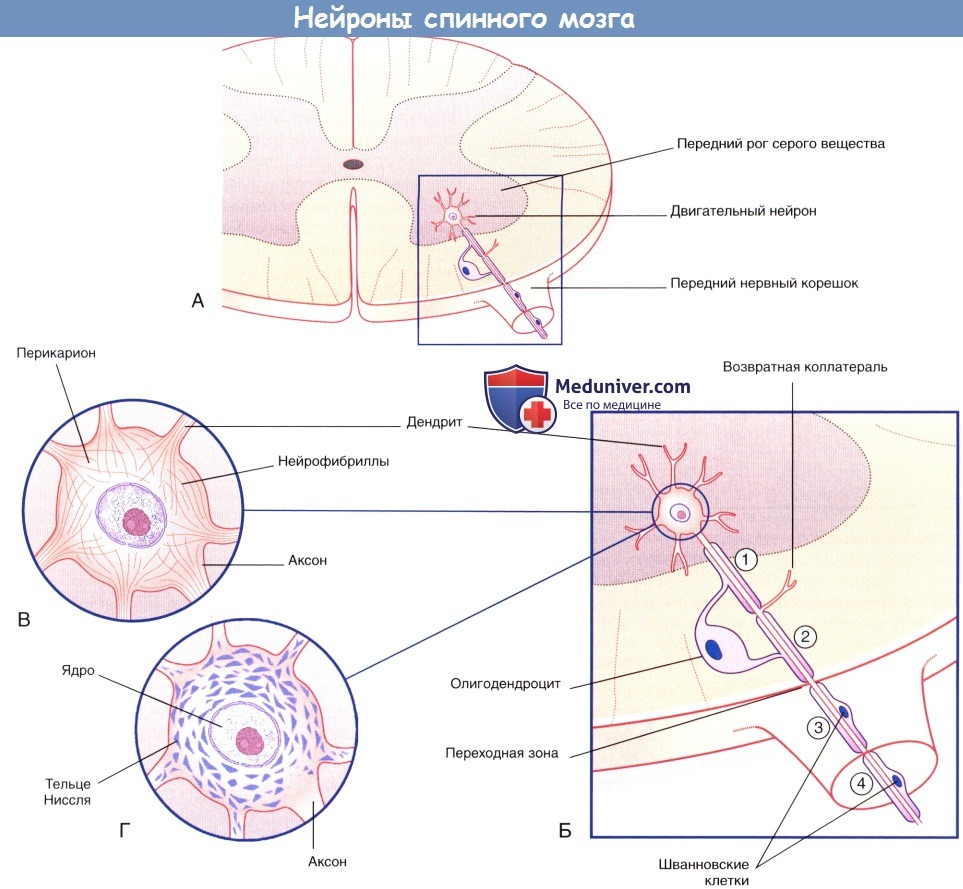 (А) Двигательный нейрон переднего рога серого вещества спинного мозга.
(А) Двигательный нейрон переднего рога серого вещества спинного мозга.
(Б) Увеличенное изображение (А). Миелиновые оболочки участков 1 и 2, располагающихся в белом веществе ЦНС, образованы олигодендроцитами.
Возвратная коллатеральная ветвь аксона начинается от немиелинизированного участка.
Миелиновые оболочки участков 3 и 4, относящихся к периферической части нервной системы, образованы шванновскими клетками.
Утолщение аксона в области вхождения в спинной мозг (переходного участка) соприкасается с одной стороны с олигодендроцитом, а с другой—со шванновской клеткой.
(В) Нейрофибриллы, состоящие из нейрофиламентов, видны после окрашивания солями серебра.
(Г) Тельца Ниссля (глыбки гранулярной эндоплазматической сети) видны при окрашивании катионными красителями (например, тионином).
Внутреннее строение нейронов
Цитоскелет всех структур нейрона образован микротрубочками и нейрофиламентами. Тело нейрона содержит ядро и окружающую его цитоплазму— перикарион (греч.peri— вокруг и karyon—ядро). В перикарионе расположены цистерны гранулярной (шероховатой) эндоплазматической сети — тельца Ниссля, а также комплекс Гольджи, свободные рибосомы, митохондрии и агранулярная (гладкая) эндоплазматическая сеть.
1. Внутриклеточный транспорт. В нейронах происходит обмен веществ между мембранными структурами и компонентами цитоскелета: непрерывно синтезируемые в соме новые клеточные компоненты перемещаются в аксоны и дендриты путем антероградного транспорта, а продукты метаболизма поступают путем ретроградного транспорта в сому, где происходит их лизосомальное разрушение (распознавание клеток-мишеней).
Выделяют быстрый и медленный антероградный транспорт. Быстрый транспорт (300-400 мм в сутки) осуществляют свободные клеточные элементы: синаптические пузырьки, медиаторы (или их предшественники), митохондрии, а также липидные и белковые молекулы (в том числе и белки-рецепторы), погруженные в плазматическую мембрану клетки. Медленный транспорт (5-10 мм в сутки) обеспечивают компоненты цнто-скелета и растворимые белки, в том числе и некоторые белки, задействованные в процессе высвобождения медиаторов в нервных окончаниях.
Аксон формирует множество микротрубочек: они начинаются от сомы короткими пучками, которые продвигаются вперед относительно друг друга вдоль начального сегмента аксона; в дальнейшем аксон формируется за счет элонгации (до 1 мм однократно). Процесс элонгации происходит за счет присоединения тубулиновых полимеров на дистальном конце и частичной деполимеризации («разборки») на проксимальном конце. В дистальной части продвижение нейрофиламентов практически полностью замедляется: в этом участке происходит процесс их достраивания за счет присоединения филаментных полимеров, поступающих в этот отдел из сомы посредством медленного транспорта.
Ретроградный транспорт метаболитов митохондрий, агранулярной эндоплазматической сети и плазматической мембраны с расположенными в ней рецепторами осуществляется с достаточно высокой скоростью (150-200 мм в сутки). Помимо выведения продуктов клеточного метаболизма, ретроградный транспорт участвует в процессе распознавания клеток-мишеней. В синапсе аксоны захватывают с поверхности плазматической мембраны клетки-мишени сигнальные эндосомы, содержащие белки,— нейротрофины («пища для нейронов»). Затем нейротрофины транспортируются в сому, где встраиваются в комплекс Гольджи.
Кроме того, захват таких «маркерных» молекул клеток-мишеней играет важную роль в распознавании клеток в процессе их развития. В дальнейшем этот процесс обеспечивает выживание нейронов, поскольку со временем их объем уменьшается, что может привести к гибели клеток в случае разрыва аксона вблизи его первых ответвлений.
Первым среди нейротрофинов был изучен фактор роста нервов, выполняющий особенно важные функции в развитии периферической чувствительной и вегетативной нервной системы. В соме нейронов зрелого мозга синтезируется фактор роста, выделенный из головного мозга (BDNF), который транспортируется антероградно в их нервные окончания. Согласно данным, полученным в результате исследований на животных, фактор роста, выделенный из головного мозга, обеспечивает жизнедеятельность нейронов, принимая участие в обмене веществ, проведении импульсов и синаптической передаче.
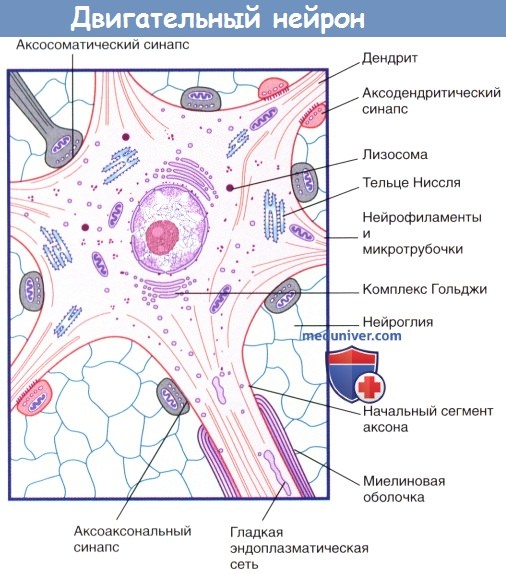 Внутреннее строение двигательного нейрона.
Внутреннее строение двигательного нейрона.
Изображены пять дендритных стволов, три возбуждающих синапса (выделены красным цветом) и пять тормозных синапсов.
2. Механизмы транспорта. В процессе нейронального транспорта роль поддерживающих структур выполняют микротрубочки. Связанные с микротрубочками белки перемещают органеллы и молекулы вдоль внешней поверхности миктротрубочек за счет энергии АТФ. Антероградный и ретроградный транспорт обеспечивают разные виды АТФаз. Ретроградный транспорт осуществляется за счет динеиновых АТФаз. Нарушение функционирования динеинов приводит к болезни двигательного нейрона.
Ниже описано клиническое значение нейронального транспорта.
Столбняк. При загрязнении раны почвой возможно заражение столбнячной палочкой (Clostridium tetani). Этот микроорганизм продуцирует токсин, который связывается с плазматическими мембранами нервных окончаний, проникает путем эндоцитоза в клетки и посредством ретроградного транспорта попадает в нейроны спинного мозга. Нейроны, расположенные на более высоких уровнях, также захватывают этот токсин путем эндоцитоза. Среди этих клеток необходимо особенно отметить клетки Реншоу, которые в норме оказывают тормозное действие на двигательные нейроны путем выделения тормозного медиатора—глицина.
При поглощении клетками токсина выделение глицина нарушается, вследствие чего прекращаются тормозные влияния на нейроны, осуществляющие двигательную иннервацию мышц лица, челюстей и позвоночника. Клинически это проявляется длительными и изнурительными спазмами этих мышц и в половине случаев заканчивается гибелью пациентов от истощения в течение нескольких дней. Предотвратить столбняк возможно, проведя своевременную иммунизацию в должном объеме.
Вирусы и токсичные металлы. Считают, что за счет ретроградного аксонального транспорта происходит распространение вирусов (например, вируса простого герпеса) из носоглотки в ЦНС, а также перенос токсичных металлов—алюминия и свинца. В частности, распространение вирусов по структурам мозга осуществляется за счет ретроградного межнейронального переноса.
Периферические нейропатии. Нарушение антероградного транспорта — одна из причин дистальных аксональных нейропатий, при которых развивается прогрессирующая атрофия дистальных участков длинных периферических нервов.
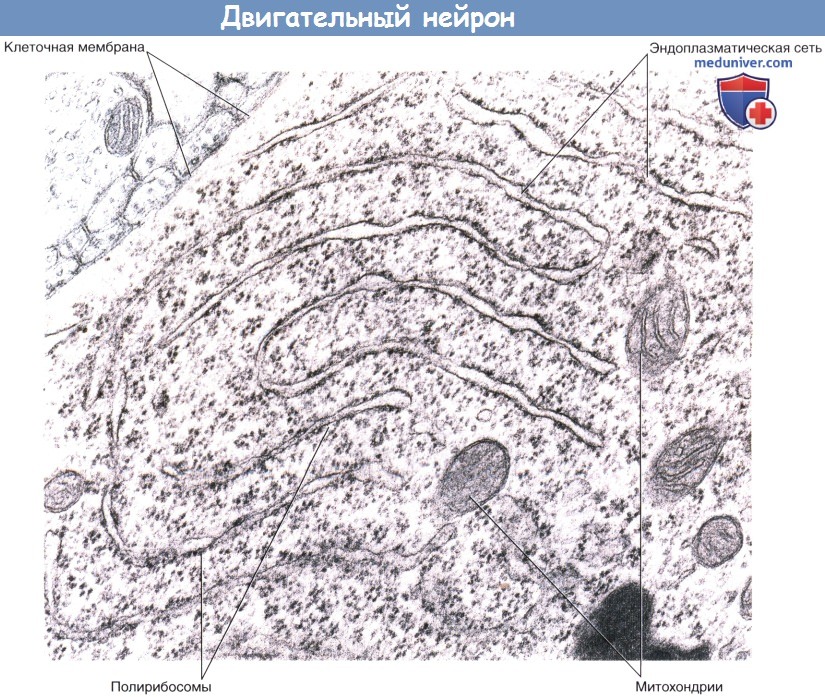 Тельце Ниссля в соме двигательного нейрона.
Тельце Ниссля в соме двигательного нейрона.
Эндоплазматическая сеть имеет многоуровневую структуру. Полирибосомы образуют выросты на внешних поверхностях цистерн или свободно лежат в цитоплазме.
(Примечание: для лучшей визуализации структуры слабо окрашены).
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Cтроение и функции нейрона
Строение и функции нейрона [ править | править код ]
Возбудимые клетки реагируют на раздражители путем изменения состояния мембран. Существуют два типа возбудимых клеток: нервные клетки, которые проводят и преобразуют импульсы в нервной системе, и мышечные клетки, которые сокращаются либо в ответ на нервные импульсы, либо автономно.
Везикулы, содержащие различные вещества (белки, липиды, сахара и молекулы медиаторов), транспортируются от комплекса Гольджи в соме к синаптическому окончанию и к кончикам дендритов путем быстрого аксонного транспорта (40 см/сутки). Этот вид антероградного (направленного вперед) транспорта по ходу нейротрубочек осуществляется кинезином (миозино-подобным белком), а энергия, необходимая для этого, поставляется АТФ. Эндогенные и экзогенные вещества, такие как фактор роста нервов (ФРН, или NRF), вирус герпеса, вирус полиомиелита и столбнячный токсин, проводятся ретроградным (направленным назад) транспортом от периферических участков к соме со скоростью
25 см/сутки. Медленный аксонный транспорт (
1 мм/сутки) играет важную роль при лечении тяжелых невритов.
Плазматическая мембрана сомы продолжается вдоль аксона и называется аксолеммой (А1, 2).
Синапс (А3) — это участок, где аксон нейрона взаимодействует с эффекторами или другими нейронами. Синаптическая передача почти у всех млекопитающих осуществляется с помощью химических соединений, а не с помощью электрических сигналов. В ответ на электрический сигнал в аксоне из везикул на пресинаптической мембране происходит высвобождение нейромедиаторов путем экзоцитоза. Медиатор диффундирует через синаптическую щель (10-40 нм) к постсинаптической мембране, где он соединяется с рецепторами, создающими новые электрические сигналы (АЗ). В зависимости от типа участвующих в процессе нейромедиатора и рецептора нейромедиатор оказывает на постсинаптическую мембрану или возбуждающий (например, ацетилхолин в скелетной мышце), или тормозящий эффект (например, глицин в ЦНС). Поскольку постсинаптическая мембрана в норме не высвобождает нейромедиаторы (существует всего несколько исключений), нервные импульсы могут пройти через синапс только в одном направлении. Таким образом, синапс действует как клапан, который обеспечивает упорядоченную передачу сигнала. Синапсы являются также участками, в которых передача нервного импульса может быть преобразована другими (возбуждающими или тормозными) нейронами.
Искусственная стимуляция нервной клетки [ править | править код ]
Когда электрический импульс из внешнего источника приложен к нервной клетке, ток течет от положительно заряженного электрода (анода) и выходит на отрицательно заряженный электрод [катод). Нервное волокно ниже катода деполяризуется, и при условии, что достигнут пороговый потенциал, генерируется потенциал действия.
Скорость проведения импульса по нерву можно измерить, поместив два электрода на кожу по ходу нерва на известном расстоянии друг от друга, с последующей стимуляцией этого нерва (содержащего многочисленные нейроны) и регистрацией времени, которое потребовалось суммарному потенциалу действия для прохождения расстояния между электродами. Скорость проведения сигнала у человека обычно составляет от 40 до 70 м/с. Значения ниже 40 м/с считаются патологическими.
Постоянный ток обычно действует как стимул только при включении и выключении: высокочастотный переменный ток (> 15 кГц), напротив, не способен вызвать деполяризацию, но повреждает ткани организма. На этом принципе основана диатермия.
Нарушения метаболизма. Биохимия специализированных тканей. (Для клин.ордов КЛД)
Введение в нейробиологию

1. Эволюция нервной системы

Чарльз Дарвин представил нам отличную теорию о зарождении и развитии жизни на Земле. Правда, в ней еще очень много вопросов и спорных моментов. Но на данном этапе лучшего у нас пока нет. Самая большая проблема с теорией мистера Дарвина в том, что мы можем догадываться и строить предположения как и почему все произошло, но мы не можем проверить или опровергнуть эту теорию. Предлагаю использовать теорию эволюции как паттерн, чтобы представить, как развивалась нервная система и какие процессы привели к появлению сознания. Мы обсудим возможные перспективы такого подхода позже.
Разнообразные типы клеток объединяются в ансамбли и паттерны, так что специализированные компоненты интегрируются в физиологию всего органа
Закроем глаза и будем представлять.
У нас есть целый океан одиночных клеток, которые переносятся потоками или плавают в лужах. Днем солнце нагревает их, а ночью они остывают. Мы можем представить, что в какой-то момент клетки научились сокращаться и этим приобрели способность к передвижению, что в совокупности с умением распознавать температуру дало им возможность перемещаться, а со временем, сокращая только одну сторону начало получаться перемещаться в более комфортном направлении.

Но некоторые все равно перемещались в неудачном направлении и погибали, и их мертвые тела, разлагаясь, создавали химический маркер для таких мест, и другие особи научились со временем различать такие маркеры. Так же со временем возможность сенсоров по распознаванию маркеров росла, и мы можем видеть, что со временем они научились распознавать не только тепло/холод, но и опасные места, где ощущаются маркеры разложения, и места где есть еда. Предлагаю называть такую систему внешними маркерами.
Система внешних маркеров, по сути, очень проста, когда на внешнем сенсоре есть определенный раздражитель, то сокращается определенная часть тела, чтобы обеспечить движение к или от раздражителя. Можно представить это как двигатель с выключателем, где роль переключателя играет внешний раздражитель. Для примера, муравей переносит упавший лист за черенок, потому что на черенке присутствует специальный химический маркер, сделанный самой природой, муравей натыкаясь на него, запускает алгоритм перетаскивания листика.

Наблюдая за насекомыми, мы можем видеть, что следующим шагом развития стала система внешних маркеров на заказ. В какой-то момент времени, мы можем предположить что внешние сенсоры научились различать не только химию разложения, но также и химические маркеры живых особей, и природа не упустила возможности создать органы способные производить различные выделения для пометок окружающей среды. Это тоже очень простая система. Например, при наличии определенного стимула в качестве переключателя животное может услышать шум и испугаться. Оно оставляет след в виде мочи или кала как маркер для других, что это опасное место.Это очень похоже на примитивную систему коммуникации. Мы можем предположить, что изначально это произошло в тот момент, когда особи научились различать свой след и возвращаться по нему назад в зону комфорта, или другие особи могли следовать за первой. Может по ночам им было более комфортно находиться вместе. Систему с внешними маркерами на заказ мы можем видеть почти у всех живых существ. Правда, у людей это чаще заметно например в таком бытовом мероприятии как покупки. Когда женская особь оставила химические маркеры на куске бумаги, а мужская особь видит совпадающие маркеры на полке, берет и кладет в корзину, вычеркивает в списке. 🙂

Но с ростом размера тел, эволюция столкнулась с парой проблем. Так как количество клеток в одной особи росло, то росло и количество сенсоров которое приходилось обрабатывать. И на этом пути эволюция собрала максимальное количество цепей обработки сигналов в одном месте. К тому же все информационно тяжелые сенсоры, зрение, слух, обоняние разместились не очень далеко.
Другой проблемой стало то, что появилась необходимость переключать весь набор клеток в различные режимы работы одновременно. Например, режимы сна, спокойствия, паники, агрессии. Реализовать это при помощи подведения командных нервов к каждой клетке не представляется возможным. И в данном случае эволюция не стала изобретать ничего нового, а просто использовала старый трюк с маркерами. Только теперь это стали маркеры внутри организма. То есть появились механизмы, начавшие вырабатывать определенные маркеры, которые разносятся кровью по организму. Клетки, раздражаясь на маркер, изменяют свой режим работы. Назовем это сигнальной системой. Самый простой и известный всем пример работы сигнальной системы, это приступ паники. Когда при наличии определенных раздражителей система начинает вырабатывать адреналин.

И вот тут мы подходим к самому интересному. На каком-то неизвестном этапе, случилось так, что некоторые особи утратили возможность распознавать внешние маркеры. У китообразных, сирен, большинства рукокрылых и узконосых приматов, включая человека, вомероназальный орган рудиментарен или полностью утрачен. На самом деле, мы, конечно, не можем знать, что случилось, было ли наращивание маркеров в сигнальной системе первичным и система внешних маркеров стала не нужна. Или сначала по какой-то причине система внешних маркеров стала недоступна и тогда началось наращивание маркеров во внутренней сигнальной системе. В случае с человеком, на данный момент известно более 100 внутренних маркеров. Так же известно, что гормоны человека могут воздействовать на животных. Но гормоны животных либо не имеют воздействия на человека, либо их реакция очень ослаблена. То есть мы можем предположить, что сигнальная система человека гораздо более развита, чем у животных. И возможно это именно та грань, которая нас различает.
Но вернемся к теме возникновения сознания.
На данном этапе наших теоретических исследований, мы пришли к тому, что у нас есть система выключателей, которые при раздражении производят какую-то работу. При этом все эти выключатели собраны физически в одном месте и крайние из них имеют в качестве подключения сенсоры или нервы.
Еще у нас имеется сигнальная система, которая управляется теми же самыми выключателями. Но при этом сами выключатели умеют реагировать на маркеры сигнальной системы. То есть мы можем это себе представить как замкнутый круг. Выключатели управляют сигнальной системой, сигнальная система воздействует на выключатели.
Имея 100 маркеров в сигнальной системе, это дает нам 2 100 вариантов коктейлей из маркеров, что дает нам огромное количество вариантов для записи сообщений между клетками. Давайте представим, что группа из выключателей при своем создании запоминает коктейль маркеров существовавший в тот момент и в дальнейшем при возникновении похожего коктейля, эта группа будет становится активной. Это обеспечивает нам связь между различными физически удаленными группами, и активацию групп при похожих ситуациях. Также не стоит забывать что производством маркеров управляют группы выключателей. Для примера, я буду очень утрировать, если мы хотим отработать блок от удара. Физически зрительная область которая отвечает за распознавание удара находится далеко ( в рамках расстояний между клетками) от области управления мышцами. Предположим что на первом этапе сообщение между этими двумя группами выключателей произойдет на уровне сигнальной системы. То есть зрительная область закажет сообщение в “сигнальной системе” в котором будет сообщение для двигательной системы произвести какие то действия. Все это будет занимать какое то время. Но вполне логично, что если повторять одно и то же действие в течении нескольких лет, то системы выключателей посылающих и принимающих сообщения отшлифуют это действие до идеала, избавят “сигнальную систему” от лишних маркеров порождающих вариативность действий, и оно перейдет почти на уровень рефлексов.

Обобщив сказанное, мы можем сказать что сознание представляет из себя бесконечный замкнутый цикл из двух систем, воздействующих друг на друга, триггеров и гормональной, в попытках найти равновесие. При этом внешние раздражители вносят свои поправки и нарушают равновесие. Эмоции в данном случае есть не более чем побочный эффект работы этих систем.
Для подтверждения данной модели, нам понадобится обнаружить нейроны которые взаимодействуют со всем спектром гормонов, что представляется мало возможным на данном уровне развития техники. Подобные нейроны исчисляются несколькими тысячами на каждый гормон, в общей массе почти сотни миллиардов нейронов.
В следующей части мы рассмотрим детальнее как из набора триггеров и сигнальной сети организовать то что мы называем сознанием. В качестве примера будем использовать реализованную компьютерную модель на ранней стадии обучения



