Водная среда заставила млекопитающих стать большими. Но не слишком

«Histoire naturelle de Lacépède» Jouvet et cie / biodiversitylibrary.org
Гигантский размер китов и других морских млекопитающих принято объяснять влиянием водной среды, однако теперь ученые обнаружили, что размер млекопитающих в воде ограничен даже сильнее, чем на суше. Исследователи выяснили, что минимальный размер водного млекопитающего ограничен необходимостью сохранять тепло, а максимальный — трудностью в получении достаточного для выживания количества пищи, говорится в статье, опубликованной в Proceedings of the National Academy of Sciences. Моделирование показало, что оптимальный вес водного млекопитающего составляет примерно 500 килограммов.
Большинство млекопитающих живут на суше, но значительная часть из них всю жизнь или большую ее часть проводят в воде. В категорию водных входят представители разных групп: отряд сирены (Sirenia), китообразные (Cetacea), отряд хищные (Carnivora) — группа Ластоногие (ушастые тюлени, настоящие тюлени, моржовые) и семейство куньи (Mustelidae). Тюлени и моржи тесно связаны с другими псообразными (Caniformia) — медведями и волками; сирены — родственники слонов; выдры и каланы близки другим куньим; а киты и дельфины приходятся родственниками бегемоту и другим парнокопытным (Artiodactyla).

Field book of giant fishes New York,G. P. Putnam / biodiversitylibrary.org
С одной стороны, большая масса помогает млекопитающим эффективнее удерживать тепло в холодной воде. С другой стороны, в воде сложнее добывать большое количество пищи активной охотой. Это затрудняет существование чересчур больших животных, поэтому водная среда практически не оставляет млекопитающим возможности быть слишком маленькими или слишком большими.
Исключением стала четвертая группа — семейство куньи (Mustelidae). Ее представители — каланы и выдры — сменили среду обитания совсем недавно и пока сохраняют небольшую массу. Возможно, это связано с тем что, они ведут полуводный образ жизни и удачно заняли свободную нишу. Исключением в большом весе стали усатые киты (подотряд Mysticeti). Эти животные — фильтраторы, и они тратят гораздо меньше энергии на добывание пищи, чем активные охотники. Поэтому усатые киты могут набирать значительно большую массу, чем их родственники, зубатые киты (подотряд Odontoceti).
Несмотря на более сильное воздействие гравитации на суше, самое большое существо, обитавшее на земле, было сухопутным. Динозавр Notocolossus gonzalezparejasi был длиннее синего кита и весил как самый крупный горбатый кит — 48 тонн. О том, как ушел золотой век больших млекопитающих на суше, читайте в нашем материале «Когда ленивцы были большими».
Морские млекопитающие: самые известные виды
Морские млекопитающие отличаются умением плавать на большие расстояния практически не испытывая усталости. Как правило, — это достаточно крупные животные, которые большую часть своего времени проводят в воде, в том числе кормят детенышей.
Морские млекопитающие: жизнь в воде
Существует 130 видов морских млекопитающих, объединенных в отряды, например, такие, как китообразные, ластоногие, выдры, сирены, белые медведи, поскольку большую часть своей жизни проводят среди морских льдов. Морские млекопитающие широко распространены по всему земному шару, но их распределение неоднородно и совпадает с продуктивностью океанов. Пики богатства находятся на широте около 40, на севере и на юге.
Эти животные приспособились к морской жизни с точки зрения конституции их тела, их рациона питания, их воспроизводства. Многие находятся под угрозой исчезновения из-за коммерческого промысла для получения мяса, кожи, жира.
Адаптации
Морские млекопитающие имеют ряд физиологических и анатомических особенностей для преодоления сложностей, связанных с водной жизнью. Некоторые из этих особенностей очень специфичны для каждого вида. Эти животные разработали ряд функций для эффективного передвижения: гидродинамические тела для уменьшения сопротивления; модифицированные конечности для плавания и маневрирования; хвостовые, спинные плавники для движения, балансирования. Эти животные специалисты в терморегуляции с использованием густого меха или жира, регулировки кровообращения (противоточные теплообменники).
Морские млекопитающие способны нырять на длительное время. Ластоногие, китообразные имеют большие и сложные системы кровеносных сосудов, которые служат для хранения кислорода необходимого при глубоководном погружении. Другие важные резервуары: мышцы, кровь, селезенка удерживают высокую концентрацию кислорода. Они также способны к брадикардии (снижение частоты сердечных сокращений) и вазоконстрикции (шунтирование большей части кислорода жизненно важным органам, таким как мозг, сердце), что позволяет увеличить время погружения и справиться с недостатком кислорода.

Существующие киты делятся на четыре семейства. Это самые большие представители отряда китообразных и самые крупные животные в мире. Особенность этих морских млекопитающих «китовый ус» — ряд роговых пластинок использующиеся для отцеживания из воды планктона, мелкой рыбы и ракообразных. Во время сна половина мозга китов остается активной, эти животные постоянно на чеку.
Для подъема на поверхность, если, например, нужно глотнуть воздуха, киты используют свой горизонтальный хвост. Эти гиганты могут пребывать под водой без подъема к поверхности около часа.
Дельфин

Также представители отряда китообразных. Дельфины – самые популярные и умные морские млекопитающие в мире. Они могут достигать девяти метров в длину, имеют вытянутую морду, питаются рыбой. Для дыхания у дельфинов на голове имеется отверстие. Чрезвычайно общительные млекопитающие, которые используют для взаимосвязи звуки, прыжки, танцы.
Ламантин

Это растительноядное животное, которое большую часть дня проводит в поисках пищи. Питается растениями в водах у берегов Африки и Америки. Единственный хищник, угрожающий ламантинам – человек. Эти гиганты могут доходить до шести метров в длину и весить 500 кг. Ламантины серого цвета, имеет уплощенный хвост и размножаются не чаще чем один раз в 2-3 года.
Морская свинья

Это китообразные относительно небольшого размера, но при этом довольно крепкие. К сожалению, данные морские млекопитающие находится под угрозой исчезновения. У этих животных округлая морда (кажется, что они всегда улыбаются), уплощенные зубы и небольшой треугольный спинной плавник.
Касатка

Млекопитающее также известное как косатка. Это еще одно очень популярное морское млекопитающее. Касатки, как и дельфины постоянные обитатели океанариумов и парков развлечений. В дикой природе обитает во всех океанах мира, хотя предпочитает умеренные и холодные зоны.
Это полуводное ластоногое млекопитающее, которое обитает в арктических морях и делится на два подвида в зависимости от района обитания: Атлантический и Тихий. Последние немного крупнее и могут весить 1 200 кг, имеют гладкую кожу толщиной в четыре сантиметра. Самцы имеют два больших клыка длиной до одного метра, а протяжность их жизни составляет около 40 лет.
Морская выдра

Этот хищник живет в северной части Тихого океана, между Японией и Мексикой. Хотя эти животные довольно часто выходят на материк большую часть своего времени они проводят в воде. Взрослые особи весят до 45 кг, имеют толстую шерсть, которая защищает их от холода. Выдры – самые маленькие морские млекопитающие. Они питаются рыбой, моллюсками, морскими ежами и ракообразными. Выдры используют камни, чтобы открывать раковины с добычей.
Тюлень
Всего существует 19 видов тюленей, которые населяют практически все прибрежные районы, кроме тропических. Тюлени мигрируют и путешествуют группами по 100 экземпляров. Задние конечности направлены назад, тюлени пользуются ими для плавания, по земле они ползают. Мех тюленей короткий, а для защиты от холода у них есть слой подкожного жира.
Эволюционная история китообразных: морское путешествие продолжительностью 55 миллионов лет
Об авторе

Покорив 375 млн лет назад сушу, позвоночные неоднократно возвращались в море, осваивая неисчерпаемые запасы рыбы и других морских организмов. Некоторые из них, например морские черепахи и ластоногие, сохранили связь с сушей, другие же полностью перешли к водному образу жизни. Хорошо известны мезозойские морские ящеры — плезиозавры, плиозавры и мозазавры. Самые большие из них достигали почти 20 м в длину. По-видимому, они охотились на своих более мелких собратьев. Хищниками были и мезозойские морские крокодилы — искусные пловцы, имевшие плавники. Но самыми успешными подводными охотниками стали ихтиозавры и китообразные, которые независимо друг от друга в совершенстве приспособились к обитанию в открытых морских водах, уподобившись рыбам формой тела и научившись размножаться без выхода на сушу.
Переход к жизни в воде сопровождался сходными преобразованиями в строении тела. Его форма становилась либо обтекаемой, либо змеевидной, уплощенной с боков. Если основным органом движения становился хвост, он приобретал вид лопасти, а задние конечности сильно уменьшались или исчезали. Передние конечности превращались в ласты или плавники, служащие веслами, рулями глубины, средствами поворота и торможения. Зубные ряды составляли двойной острый частокол, способный удерживать и раздирать скользкую добычу.
После вымирания гигантских водных рептилий мезозоя ниша морских суперхищников долго оставалась свободной и лишь частично осваивалась большими акулами, крокодилами и морскими змеями. Но около 55 млн лет назад появилась группа млекопитающих, со временем достигшая господства в морях кайнозойской эры, — китообразные.
Родственные узы
Китообразные — потомки наземных млекопитающих, и наследие их сухопутного прошлого проявляется в дыхании атмосферным воздухом, наличии плаценты, вскармливании детенышей молоком, а также в деталях строения скелета. В отличие от рыб, у которых при плавании позвоночник изгибается в горизонтальной плоскости, у китообразных он двигается в плоскости вертикальной. Современные киты и дельфины не могут жить на суше и выглядят очень необычно по сравнению с наземными млекопитающими. Вопрос о том, каким образом наземные звери превратились в морских чудовищ, долгое время оставался без ответа, так как в палеонтологической летописи отсутствовали соответствующие связующие звенья. В настоящее время картина в значительной мере прояснилась.
Предками китообразных до 2000-х годов считались мезонихии, родственные парнокопытным вымершие хищники, обликом отдаленно напоминавшие массивных собак с копытцами вместо когтей. Мезонихии были величиной от ласки до медведя, среди них известны и полуводные рыбоядные формы, такие как гапалодекты (Hapalodectes). Однако большинство мезонихий были наземными хищниками и падальщиками. Строение черепа и зубов этих древних млекопитающих (они жили 63–33 млн лет назад) долгое время давало основание полагать, что китообразные произошли непосредственно от мезонихий.
С этой гипотезой вступили в противоречие молекулярно-генетические данные, показывающие, что китообразные должны рассматриваться в пределах группы парнокопытных в качестве ближайших родственников бегемотов. На этом основании парнокопытные и китообразные иногда объединяются в группу «китопарнокопытных» (Cetartiodactyla) [1].
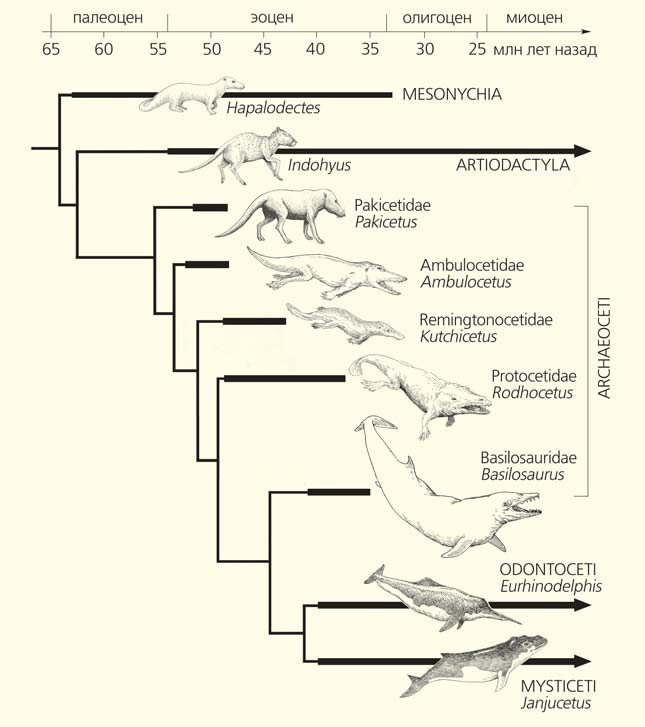
Схема эволюции китообразных. Схема А. В. Лопатина, рисунки животных К. К. Тарасенко
Открытие полного скелета наземного китообразного пакицета (Pakicetus attocki) поддержало молекулярно-генетические данные, показав, что китообразные отделились от древних парнокопытных уже после того, как те разошлись с мезонихиями [2]. Иными словами, предками китов были примитивные парнокопытные, сохранившие признаки их общего с мезонихиями происхождения (например, в строении зубов), которые современные парнокопытные давно утратили. Это были всеядные и плотоядные животные. Парнокопытные позже перешли к питанию растительным кормом, но некоторые из них (например, свиньи) и ныне в значительном объеме потребляют животную пищу. Киты — водные теплокровные, нуждающиеся в высококалорийной пище, поэтому они сохранили и развили животноядный характер питания своих предков.
В пользу этой гипотезы может свидетельствовать строение скелета индохиуса (Indohyus), который был найден в Кашмире (Индия) в отложениях возрастом около 48 млн лет [3]. Это животное относится к отряду парнокопытных, к вымершему семейству раоэллид. Телосложением и величиной оно напоминало вовсе не китов, а современных оленьков семейства трагулид, живущих в Западной Африке и Юго-Восточной Азии в густых лесах, кустарниковых и мангровых зарослях, умеющих хорошо плавать и нырять. Но по строению слуховой области черепа (это очень важный признак для выяснения родства среди млекопитающих) индохиус сближается лишь с одними древнейшими китами семейства пакицетид. Индохиус жил на несколько миллионов лет позже пакицетид и не может быть их предком — но он вполне может быть потомком одной из линий, родственной предкам династии морских великанов.
Основатели династии
Древние китообразные эволюционировали быстро, и переход от полуназемных форм к постоянноводным занял приблизительно 8 млн лет. Примитивных эоценовых китообразных, называемых археоцетами (древними китами), разделяют на пять семейств — это пакицетиды, амбулоцетиды, ремингтоноцетиды, протоцетиды и базилозавриды. В 2001 г. стали известны первые анатомически полные скелеты археоцетов, позволившие в деталях выяснить процесс перехода китообразных к жизни в море [2].
Пакицетиды жили в раннем эоцене (около 52 млн лет назад) на северном побережье субтропического внутриконтинентального моря Тетис, в те времена пересекавшего Евразию в широтном направлении. Это были большеголовые и длинномордые звери с копытцами на пальцах и длинными хвостами, пакицет и налацет (Nalacetus ratimitus) размерами соответствовали волку, а ихтиолест (Ichthyolestes pinfoldi) — лисице. Судя по строению скелета, пакицетиды хорошо плавали, используя волнообразные движения тела и гребки конечностями. По строению слуховой области черепа они сближаются с китами. Однако уши пакицетид были приспособлены к восприятию звуков в воздушной, а не в водной среде. Следовательно, эти животные жили на суше, добывая пищу в мелководных пресных водоемах и поблизости от них, подобно современным тапирам. Скелеты пакицетид были найдены в Пакистане и Северо-Западной Индии, в речных отложениях. Видимо, именно эти места были центром происхождения китообразных.
Данные по эволюции системы полукружных каналов внутреннего уха, участвующих в регуляции равновесия и положения тела в пространстве и ответственных за контроль локомоции, свидетельствуют, что пакицетиды в этом отношении еще были близки наземным млекопитающим [4].
Киты-амфибии
Одной из самых значимых находок древних китообразных, «недостающим звеном» в их эволюции, стал амбулоцет (Ambulocetus natans). Его название, означающее «плавающий ходячий кит», говорит само за себя. Почти полный скелет этого животного найден на севере Пакистана в отложениях возрастом около 49 млн лет назад [5]. Обликом трехметровый амбулоцет напоминал большеголового крокодила. При плавании его тело изгибалось в вертикальной плоскости, как у современных китов, тюленей и выдр. Предполагают, что амбулоцеты охотились подобно крокодилам, подстерегая своих жертв на мелководье (активно передвигаться по суше они не могли). Амбулоцеты имели мощные челюсти и зубы и были способны умертвить достаточно крупную добычу. Строение носа позволяло «киту-амфибии» глотать пищу прямо в воде. Глаза давали только боковой обзор. Ушные раковины амбулоцет, судя по всему, утратил, но хорошо слышал в воде. Движение своей добычи по суше он отслеживал, прижимая голову к грунту и улавливая колебания субстрата. Химический анализ зубов показал, что зверь мог охотиться как в солоноводных, так и в пресных водоемах. К семейству амбулоцетид относятся еще два рода: гандаказия (Gandakasia) и гималайяцет (Himalayacetus). Последний примечателен своей древностью, наибольшей среди китообразных — 53,5 млн лет.

Реконструкция внешнего вида амбулоцета. Рисунок К. К. Тарасенко
Более мелкими родичами амбулоцетид были ремингтоноцетиды, известные из Индии и Пакистана, из отложений возрастом 49–43 млн лет назад. Эти животные обладали сильно вытянутыми челюстями и были несколько лучше приспособлены к подводному плаванию, чем амбулоцетиды. Они тоже имели хорошо развитые конечности, но отличались маленькими глазами, тонкими челюстями и расширенным основанием черепа. Если сравнивать «земноводных китов» по общему облику с крокодилами, то среди них амбулоцетиды больше напоминали аллигаторов, а ремингтоноцетиды — гавиалов. Строение слуховой области в сочетании с маленькими глазами показывает, что для обнаружения добычи ремингтоноцетиды пользовались в основном слухом. Известно шесть родов ремингтоноцетид. Их размеры были невелики: например, качхицет (Kutchicetus minimus) не превосходил величиной речную выдру. Для эндрюсифия (Andrewsiphius sloani) установлено наличие мощного уплощенного хвоста, служившего основным органом движения в водной среде [6]. Райанист (Rayanistes afer), найденный в среднем эоцене Египта [7], демонстрирует, что ремингтоноцетиды достигли южного побережья Тетиса.
Киты дальнего плавания
Первыми с мелководья на настоящую глубину отправились протоцетиды, жившие 49–37 млн лет назад. Их главным эволюционным приобретением был удлиненный мощный хвост (поначалу без хвостовой лопасти), который обеспечивал быстрое плавание [8]. При этом протоцетиды сохраняли развитые задние конечности. Вероятнее всего, по образу жизни их можно сопоставить с современными крупными ластоногими. У некоторых протоцетид зубы были уменьшены, это свидетельствует, что добычу — мелких морских животных — они заглатывали целиком. Такими были первые китообразные, которые сумели распространиться за пределы моря Тетис — их остатки найдены в мелководно-морских отложениях в Африке, Европе и Северной Америке. Самые крупные из них достигали длины более 3 м и весили до 400–500 кг. Вероятно, от протоцетид произошли все более поздние китообразные, включая современные группы. Описано около 20 родов протоцетид, из них наиболее известны протоцет (Protocetus) и родхоцет (Rodhocetus). Своеобразно выглядел макарацет (Makaracetus bidens), у которого (судя по строению передней части черепа) был короткий хоботок [9] или мясистая верхняя губа, которые могли служить для сбора со дна моллюсков и других животных.
Важные сведения о биологии протоцетид были получены на основе изучения майяцета (Maiacetus inuus). В Пакистане, в отложениях возрастом 47,5 млн лет, был обнаружен скелет самки, внутри которого сохранились череп и часть скелета эмбриона [10]. Его положение (вперед головой, а не хвостом, т.е. так, как у наземных млекопитающих) свидетельствует, что протоцетиды рождали детенышей на суше, а не в воде. Самцы майяцетов были крупнее самок и имели более мощные клыки. Такой половой диморфизм характерен для многих современных ластоногих, в частности моржей и морских львов. Возможно, он отражает сходный, «гаремный», тип социальной организации у древних китообразных.
Властелины древних морей

Реконструкция внешнего вида базилозавра. Рисунок В. Д. Калганова
Первые действительно гигантские киты — базилозавриды — жили в позднем эоцене, приблизительно 41–35 млн лет назад. Их ископаемые остатки в основном происходят с востока США и из Египта, но, скорее всего, эти животные были распространены по всему свету [11]. Известно около 15 родов этих археоцетов. Представители подсемейства базилозаврин были огромны: их змеевидное тело массой до 6 т достигало в длину 15–20 м. У них имелся хвостовой плавник, но был ли он их основным органом движения — неизвестно. Как и для прочих древних китов, для базилозаврин были характерны зазубренные коренные зубы и конические предкоренные. Дорудонтины внешне походили на дельфинов (хотя имели заметную шею), при весе в 1,5 т достигали в длину до 6 м. Все базилозавриды имели вполне оформленные задние конечности с подвижным коленным суставом и несколькими пальцами. Однако эти конечности были совсем маленькими и явно не функционировали как органы движения. Возможно, самцы использовали их при спаривании для захватывания самок, как делают своими рудиментарными задними конечностями удавы. Несмотря на внешнее сходство с современными китообразными, базилозавриды не были способны к ультразвуковой эхолокации и акустической коммуникации. Относительно маленький мозг указывает на отсутствие у них сложного социального поведения, характерного для современных китов и дельфинов. В то же время явно выраженная асимметрия черепа базилозаврид была частью комплекса приспособлений, связанного с развитием у них направленного слуха для различения высокочастотных звуков, которые производили рыбы.
Самый крупный археоцет (и самое крупное млекопитающее своего времени) — это базилозавр (Basilosaurus cetoides). Он был описан еще в 1839 г. из Алабамы (США). Другие виды найдены в Египте и Пакистане. Строение позвоночника базилозавра позволяет предполагать, что при плавании он мог змеевидно извиваться. Скорее всего, базилозавр не был способен к длительному плаванию и глубокому нырянию. Не мог и выходить на сушу. Вероятно, он охотился на крупную добычу (например, акул, а также дорудонтин) у водной поверхности недалеко от побережья. В 2015 г. в Египте нашли полный скелет 18-метрового базилозавра с остатками другого кита внутри.

Слепок черепа Basilosaurus cetoides, вид снизу. Экспонат Палеонтологического музея имени Ю. А. Орлова, Палеонтологический институт имени А. А. Борисяка РАН. Фото А. А. Ермакова
Между зубом и усом
У современных китов задние конечности рудиментарны — они скрыты внутри тела и служат для крепления мышц половых органов и лишь поэтому не исчезли полностью. Иногда рождаются киты с атавистическими миниатюрными задними конечностями. Вместо ноздрей современные киты имеют дыхало специфичного строения, смещенное на макушку. Строение ушной системы обеспечивает способность к акустической коммуникации. Современные группы китообразных — зубатые (Odontoceti) и усатые (Mysticeti) киты — появилсь в позднем эоцене, около 40 млн лет назад. К раннему олигоцену они стали обычны, а в позднем олигоцене (27–23 млн лет назад) дали мощную вспышку разнообразия, которая привела к возникновению большинства современных семейств. Резкое увеличение головного мозга у зубатых китов отмечается дважды: первоначально у самых ранних форм 40 млн лет назад и затем у дельфинов 15 млн лет назад [12].
Подотряд зубатых китов включает дельфинов, нарвалов, морских свиней и кашалотов. Большинство из них живут в океане (хотя есть речные дельфины), известны прибрежные и океанические виды, полярные и тропические. Все они активные хищники, добывают рыбу и кальмаров или, как косатки, охотятся на других китообразных, а также на пингвинов и ластоногих. В отличие от современных усатых китов, зубатые имеют зубы и способны к эхолокации. Каждое из современных семейств подотряда обладает собственной специализацией. Не менее разнообразны были и вымершие группы.
Современные усатые киты утратили зубы, но зато приобрели китовый ус, служащий им для фильтрации больших объемов мелкой добычи. Среди усатых китов — крупнейшее создание мира животных за всю его историю — синий кит.
Некоторые необычные представители как зубатых, так и усатых китов, в прошлом населявшие моря нашей планеты, заслуживают более близкого знакомства.
Мечерылые, акулозубые, моржеголовые.

Реконструкция внешнего вида сквалодона. Рисунок И. С. Сергеенковой
Симоцет (Simocetus rayi) из верхнего олигоцена Орегона (США), выделенный в самостоятельное семейство симоцетид, получил прозвание «курносый дельфин». Его короткие челюсти были заметно загнуты вниз, а редко расположенные зубы служили скорее для просеивания пищи, чем для ее разрывания. Вероятно, симоцет добывал донных беспозвоночных, процеживая осадок [14].
У двухметровой морской свиньи семиростра (Semirostrum ceruttii), жившей 3 млн лет назад у берегов южной Калифорнии (США), нижняя челюсть была вдвое длиннее верхней. Эта часть челюсти формировала покрытый кожей выступ-подбородок, который использовался как специальный чувствительный орган для поиска пищи «на ощупь» в мутной воде у самого дна [15].
История группы речных дельфинов (Platanistoidea) прослеживается с позднего олигоцена, когда ее представители еще обитали в морских бассейнах. При этом оказывается, что они вовсе не близкородственны друг другу. Слепой гангский дельфин (Platanista) принадлежит линии архаичных зубатых китов, родственной сквалодонтидам, тогда как другие — амазонский (Inia), лаплатский (Pontoporia) и озерный (Lipotes) — скорее всего, происходят от различных вымерших групп [16].

Реконструкция внешнего вида одобеноцетопса. Рисунок И. С. Сергеенковой
Кашалоты-убийцы и сам Левиафан
В раннем миоцене, около 20 млн лет назад, появились кашалоты (Physeteroidea). Ныне это специализированные охотники на кальмаров. Однако некоторые неогеновые кашалоты были китами-убийцами своего времени, занимая экологическую нишу косаток до их появления около 3 млн лет назад. Наименование «кашалота-убийцы» в научной литературе получил зигофизетер (Zygophyseter varolai) из позднего миоцена Италии, возрастом 10–7 млн лет назад [18]. Этот кит был длиной 6–7 м, с мощным черепом и многочисленными большими зубами в верхней и нижней челюстях. Грозными хищниками были и другие кашалоты-убийцы: семиметровый бригмофизетер (Brygmophyseter shigensis) из Японии (возраст 15–14 млн лет назад) и представители рода акрофизетеров (Acrophyseter) из Перу (9–6 млн лет), достигавшие в длину лишь 4 м, но также занимавшие нишу активных морских охотников [19].
Остатки намного более крупного морского хищника были также обнаружены в Перу, их возраст 10–9 млн лет назад [19, 20]. В длину гигантский кашалот-убийца достигал 18 м, а весил он около 40 т (примерно как самцы современного кашалота). Длина его черепа около 3 м, а зубы имеют диаметр около 12 см и длину до 36 см. Пожалуй, это самые крупные зубы в животном мире, не считая гипертрофированных клыков и бивней. Колосс получил название «мелвиллов левиафан» (Livyatan melvillei), в честь библейского чудовища Левиафана и американского писателя Германа Мелвилла, автора романа «Моби Дик, или Белый кит». Можно предполагать, что появление таких крупных хищников было связано с развитием гигантизма среди их потенциальных жертв — усатых китов.
Киты-леопарды и беззубые великаны
Усатые киты произошли от археоцетов, по-видимому, в высоких широтах. Известны олигоценовые группы, которые имели широко расставленные зубы, а в строении черепа совмещали черты, свойственные археоцетам и усатым китам. Очень необычен зубастый янджуцет (Janjucetus hunderi) из позднего олигоцена Австралии, длиной около 3,5 м, имевший исключительно большие глаза и укороченную морду. Предполагается, что он был активным хищником, охотившимся на крупную рыбу, включая акул, и экологически напоминал морского леопарда — современного хищного тюленя [21]. Янджуцет относится к семейству маммалодонтид. Другие представители этой вымершей группы, маммалодоны (Mammalodon) из олигоцена Австралии и Новой Зеландии, имели редко посаженные зубы. Вероятно, они питались донными беспозвоночными, процеживая ил [22]. Их открытие подтвердило догадку Чарлза Дарвина, который предположил в «Происхождении видов», что фильтрующие усатые киты произошли именно от таких форм — Дарвин называл их «просеивателями».

Реконструкция внешнего вида янджуцета. Рисунок И. С. Сергеенковой
В середине олигоцена, около 30 млн лет назад, появились фильтрующие усатые киты. У представителей вымершего семейства эомистицетид (Eomysticetus, Waharoa и др.), сравнительно некрупных (до 8 м) китов стройного сложения, китовый ус занимал только заднюю часть неба, на переднем конце челюсти имелось несколько зубов, а дыхало располагалось на значительном расстоянии спереди от глаз. Древнейшие находки гладких китов (Balaenidae) имеют возраст 23 млн лет. Полосатиковые (Balaenopteridae), к которым относится гигантский синий кит, появились в Северной Атлантике около 12 млн лет назад. Судя по ископаемым остаткам, современному синему киту по размерам не уступали некоторые киты-полосатики плиоцена и плейстоцена, например Balaenoptera sibbaldina из раннего плиоцена Бельгии [23]. Однако самыми широко распространенными усатыми китами неогена были цетотерии (Cetotheriidae) — очень разнообразная (более 50 видов и около 20 родов) группа сравнительно небольших форм (около 3–4 м) без признаков специализации современных семейств. Судя по всему, в основном на них и охотились кашалоты-убийцы, а также гигантские акулы мегалодоны. Остатки цетотериев известны из Европы, Северной и Южной Америки, Японии, Австралии и Новой Зеландии.

Реконструкция внешнего вида цетотерия. Рисунок И. С. Сергеенковой
Итак, появившись почти 55 млн лет назад в прибрежных водах Индо-Пакистанской области, китообразные постепенно освоили Мировой океан, породив огромное множество форм весьма разнообразного облика и величины.

Скелет Vampalus sayasanicus. Вверху — процесс раскопок (видны части черепа, нижней челюсти, позвонки), внизу — фрагмент черепа: вид сверху (слева) и снизу. Длина масштабной линейки 10 см. Фото Х. А. Абдулмежидова (верхнее) и К. К. Тарасенко (нижние)
Ископаемая летопись китообразных постоянно пополняется яркими находками, сулящими дальнейшие открытия. В России остатки вымерших китов встречаются довольно редко, в основном в отложениях миоцена (Ставрополье, Дагестан, Таманский п-ов, Сахалин). В Ростовской обл. обнаружены фрагментарные остатки среднеэоценовых археоцетов [24]. На Северном Кавказе находят цетотериев. Недавно оттуда были описаны несколько новых родов этих китов. Важную находку передал в Палеонтологический институт имени А. А. Борисяка РАН житель Чеченской республики Х. А. Абдулмежидов, раскопавший в Ножай-Юртовском р-не почти полный скелет кита. Этот новый кит из группы цетотериев получил название «вампал» (Vampalus sayasanicus), по имени великана из чеченской мифологии [25].
Литература
1. Grauer D., Higgins D. Molecular evidence for the inclusion of cetaceans within the order Artiodactyla // Molecular Biology and Evolution. 1994; 11(3): 357–364.
2. Thewissen J. G. M., Williams E. M., Roe L. J., Hussain S. T. Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls // Nature. 2001; 413: 277–281.
3. Thewissen J. G. M., Copper L. N., Clementz M. T. et al. Whales originated from aquatic artiodactyls in the Eocene epoch of India // Nature. 2007; 450(7173): 1190–1194.
4. Spoor F., Bajpai S., Hussain S. T. et al. Vestibular evidence for the evolution of aquatic behaviour in early cetaceans // Nature. 2002; 417(6885): 163–166.
5. Williams E. M. Synopsis of the earliest cetaceans: Pakicetidae, Ambulocetidae, Remingtonocetidae, and Protocetidae // The emergence of whales. Evolutionary patterns in the origin of Cetacea. N.Y., 1998: 1–28.
6. Thewissen J. G. M., Bajpai S. New skeletal material of Andrewsiphius and Kutchicetus, two Eocene cetaceans from India // Journal of Paleontology. 2009; 83(3): 635–663.
7. Bebej R. M., Zalmout I. S., Abed El-Aziz A. A. et al. First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution // Journal of Paleontology. 2016; 89(5): 882–893.
8. Bajpai S., Thewissen J. G. M., Sahni A. The origin and early evolution of whales: macroevolution documented on the Indian Subcontinent // Journal of Biosciences. 2009; 34(5): 673–686.
9. Gingerich P. D., Zalmout I. S., Ul-Haq M., Bhatti M. A. Makaracetus bidens, a new protocetid archaeocete (Mammalia, Cetacea) from the Early Middle Eocene of Balochistan (Pakistan) // Contributions from the Museum of Paleontology, University of Michigan. 2005; 31(9): 197–210.
10. Gingerich P. D., Ul-Haq M., Koenigswald W. von et al. New protocetid whale from the Middle Eocene of Pakistan: birth on land, precocial development, and sexual dimorphism // PLoS One. 2009; 4(2): e4366: 1–20.
11. Thewissen J. G. M., Williams E. M. The early radiation of Cetacea (Mammalia): evolutionary pattern and developmental correlation // Annual Review of Ecology and Systematics. 2002; 33: 73–90.
12. Marino L., McShea D. W., Uhen M. D. Origin and evolution of large brains in toothed whales // Anatomical Record. Pt A. 2004; 281A: 1247–1255.
13. Geisler J. H., Colbert M. W., Carew J. L. A new fossil species supports an early origin for toothed whale echolocation // Nature. 2014; 508(7496): 383–386.
14. Fordyce R. E. Simocetus rayi (Odontoceti: Simocetidae) (new species, new genus, new family), a bizarre archaic Oligocene dolphin from the eastern North Pacific // Smithsonian Contributions to Paleobiology. 2002; 93: 185–222.
15. Racicot R. A., Deméré T. A., Beatty B. L., Boessenecker R. W. Unique feeding morphology in a new prognathous extinct porpoise from the Pliocene of California // Current Biology. 2014; 24(7): 774–779.
16. Hamilton H., Caballero S., Collins A. G., Brownell R. L. Evolution of river dolphins // Proceedings of the Royal Society of London. Ser. B. 2001; 268: 549–556.
17. Muizon C. de. Walrus-like feeding adaptation in a new cetacean from the Pliocene of Peru // Nature. 1993; 365(6448): 745–748.
18. Bianucci G., Landini W. Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy // Zoological Journal of the Linnean Society. 2006; 148: 103–131.
19. Lambert O., Bianucci G., Muizon C. de. Macroraptorial sperm whales (Cetacea, Odontoceti, Physeteroidea) from the Miocene of Peru // Zoological Journal of the Linnean Society. 2017; 179: 404–474.
20. Lambert O., Bianucci G., Post K. et al. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru // Nature. 2010; 466(7302): 105–108.
21. Fitzgerald E. M. G. A bizarre new toothed mysticete (Cetacea) from Australia and the early evolution of baleen whales // Proceedings of the Royal Society of London. Ser. B. 2006. 273(1604): 2955–2963.
22. Fordyce R. E., Marx F. G. Mysticetes baring their teeth: a new fossil whale, Mammalodon hakataramea, from the Southwest Pacific // Memoirs of Museum Victoria. 2016; 74: 107–116.
23. Deméré T. A., Berta A., McGowen M. R. The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes // Journal of Mammalian Evolution. 2005; 12(1/2): 99–143.
24. Тесаков А. С., Александрова Г. Н., Тарасенко К. К. и др. Палинологическая характеристика палеогеновых отложений и находка древнего кита на юго-западном побережье Цимлянского водохранилища (Ростовская область) // Бюллетень Региональной межведомственной стратиграфической комиссии по центру и югу Русской платформы. Вып. 5. М., 2012: 138–143.
25. Тарасенко К. К., Лопатин А. В. Новые роды усатых китов (Cetacea, Mammalia) из миоцена Северного Кавказа и Предкавказья. 2: Vampalus gen. nov. (средний — поздний миоцен, Чечня и Краснодарский край) // Палеонтол. журн. 2012; 6: 72–81.
* См. подробный обзор: Гельфанд М. С. Молекулярная эволюция: как киты уходили под воду // Природа. 2016. № 10. С. 39–50.



